

| {1} |
|
KNUT SCHMIDT-NIELSEN HOW CAMBRIDGE UNIVERSITY PRESS |
| {2} |
|
КНУТ ШМИДТ-НИЛЬСЕН КАК РАБОТАЕТ Перевод с английского Под редакцией и с предисловием ИЗДАТЕЛЬСТВО «МИР» |
| {3} |
591.4
Ш73
|
Шмидт-Нильсен К. Как работает организм животного. Пер. с англ. Н. О. Фоминой. Под ред. и с предисл. Н. Н. Карташева. М., «Мир», 1976. 144 с. (В мире науки и техники) Книга английского физиолога Кнута Шмидта-Нильсена, известного своими исследованиями в области сравнительной и экологической физиологии, посвящена механизму работы некоторых систем организма животного. Автор показывает, как использование принципа противотока позволяет организму позвоночных животных сохранять или рассеивать образующееся в нем тепло, сокращать энергетические затраты, поддерживать благоприятный водный баланс, интенсифицировать дыхание. Книга предназначена в первую очередь для студентов биологических факультетов и преподавателей биологии, однако она может оказаться полезной и специалистам — биологам и инженерам, работающим в области бионики, а также всем тем, кого просто интересует биология.
591.4 Редакция научно-популярной © Перевод на русский язык, «Мир», 1976 |
| {4} |
Как работает организм животного? Какими путями обеспечивается его приспособленность к постоянно меняющимся условиям окружающей среды? Ответы на эти вопросы стремятся найти специалисты разных отраслей биологии.
Животный мир Земли очень разнообразен: более миллиона видов населяют сейчас нашу планету. Каждый из них отличается своими морфологическими, физиологическими, экологическими, поведенческими особенностями. Эти особенности вырабатываются в процессе эволюции и определяют видовую специфику взаимоотношений животного с окружающей средой. Между близкими видами различия иногда выявляются лишь с большим трудом. Чем дальше отстоят друг от друга сравниваемые виды, тем резче они отличаются, тем больше разнятся общие принципы и конкретные механизмы их приспособлений к условиям жизни.
И тем не менее всесторонние сравнительные исследования многих видов из разных таксономических групп (а значит, и различной филогенетической близости) позволяют не только выяснить видоспецифичные особенности, обеспечивающие выживание каждого данного вида в непрерывно идущей борьбе за существование, но и выявить общие черты строения и функций, общие принципы приспособлений к окружающей среде {5} в пределах разных систематических групп. Об одних общих закономерностях строения и функционирования органов, их систем и организма в целом мы сейчас знаем достаточно детально, о других пока имеем лишь самое приближенное представление.
Относительно недавно было выяснено, что в разных группах животных при выработке приспособлений к меняющимся условиям среды довольно широко используется принцип противотока. Как этот принцип используется позвоночными животными, рассказывает предлагаемая читателю книга К. Шмидта-Нильсена.
Она представляет собой изложение лекций, прочитанных им в 1970—1971 учебном году в Кембриджском университете. На примере отдельных исследований, проведенных в последние годы как самим автором и его учениками, так и другими учеными, ярко и убедительно показана роль противоточных систем (кровь — воздух, кровь — кровь и др.) в сохранении и рассеивании образующегося в организме тепла, в сокращении энергетических затрат, в интенсификации дыхания, в поддержании благоприятного водного и солевого баланса. В заключительной главе рассматриваются некоторые физиологические проблемы, связанные с характерным для позвоночных животных очень большим диапазоном абсолютных размеров тела (например, среди млекопитающих мелкие землеройки весят 3—4 грамма, слоны 3—4 тонны, киты 20—50 и даже 100 тонн). Все эти данные достаточно новы и еще не получили всестороннего отражения не только в научно-популярной, но и в учебной литературе.
Несколько слов об авторе книги. Кнут Шмидт-Нильсен родился в 1915 году в Тронхейме, окончил школу в Норвегии, затем поступил в Копенгагенский университет, где учился у Августа Крога, а диссертацию на степень доктора философии готовил в Карлсбергской {6} лаборатории под руководством Кая Линдестрема-Ланга. В 1946 году Шмидт-Нильсен начал изучать животный мир в пустынях Северной Америки. С 1952 года он профессор физиологии на зоологическом отделении Университета Дьюка.
Физиолог по образованию, физиолог и эколог по опыту работы, строгий экспериментатор и вдумчивый полевой натуралист, профессор Кнут Шмидт-Нильсен — крупный ученый, успешно работающий в области экологической физиологии. Особенно широко известны его оригинальные работы по исследованию эколого-физиологических приспособлений животных к жизни в пустыне; они обобщены в переведенной на русский язык книге «Животные пустынь. Физиологические проблемы тепла и воды» (изд-во «Наука», 1972). Большой интерес вызвали его исследования по выяснению роли носовых и надорбитальных желез у пресмыкающихся и птиц, эффективно выводящих из организма излишки солей, по определению энергетических затрат у разных животных, новая трактовка действия дыхательной системы птиц и другие работы. Для всех исследований К. Шмидта-Нильсена характерен комплексный синтетический подход к изучаемым явлениям и объектам, стремление выявить взаимосвязи морфофизиологических особенностей с образом жизни животного, поиск общих принципов приспособления животных к условиям окружающей среды.
Стиль ученого отчетливо проявляется и в предлагаемой книге. Читатель, несомненно, оценит широкую эрудицию автора, четкость и логичность повествования, а также новизну приводимых фактов, хотя сжатость изложения потребует при чтении постоянного внимания и сосредоточенности — к разряду развлекательных эта книга не относится. {7}
Книга адресована в первую очередь студентам старших курсов, специализирующимся в разных разделах биологии (физиологам, морфологам, экологам), и преподавателям биологии в школах. Однако можно не сомневаться, что ее с удовольствием прочтут и специалисты-биологи, а также инженеры, работающие в области бионики. Излагаемые автором факты и идеи, вероятно, привлекут и всех тех, кто просто интересуется биологией.
Н. Н. Карташев
| {8} |
Заинтересовавшись какой-нибудь простенькой биологической проблемой и приступив к ее изучению, мы вдруг обнаруживаем, что по мере углубления в нее возникает множество новых вопросов, отчего проблема все более усложняется. Появляются новые проблемы, казалось бы, вовсе не связанные с первоначальной, однако в конечном счете все они сводятся к стремлению узнать, что и каким образом происходит вокруг. Если повезет, то нам удастся кое-что выяснить, а в случае настоящей удачи, когда мы постигаем принципы, лежащие в основе изучаемого явления, самой величайшей наградой оказывается простота этих принципов.
Двадцать пять лет назад мне представилась возможность посетить пустыню Аризоны. Я был поражен богатством ее животного мира, совершенно для меня неожиданным. Особенно меня удивило обилие мелких грызунов — ведь воды, которую они могли бы пить, вокруг не было. Не менее сильное впечатление произвело на меня полное отсутствие в научной литературе данных (за исключением нескольких полуанекдотических сообщений) о том, как эти зверьки все-таки умудряются жить, по-видимому, совсем без воды. Сейчас твердо установлено, что водный баланс обитающих в пустыне грызунов, например кенгуровой крысы, ничем не отличается от водного баланса любого другого животного, то есть для поддержания нормального водного баланса она, как и всякое животное, должна получить столько воды, сколько выделяет. Стало известно также, что в течение длительных периодов засухи животные пустыни вовсе не прозябают, а чувствуют себя так же хорошо, как и любое другое животное, находящееся в своей естественной среде. {9}
Рассмотрим вкратце водный баланс кенгуровых крыс, которые питаются семенами и прочей сухой растительной пищей. Они обычно не пьют воды и не выискивают сочных зеленых растений. Упрощенная картина их водного баланса, представленная в таблице 1,
Таблица 1
Водный баланс кенгуровой крысы
Для поддержания водного баланса животного
суммарное поступление воды в организм должно быть равно
ее суммарным потерям
Поступление, % |
Потери, % |
||
Вода, образующаяся в |
90 |
Испарение |
70 |
Свободная вода, содер- |
10 |
С мочой |
25 |
Питьевая вода |
0 |
С калом |
5 |
позволяет ответить на все возникающие вопросы. Поскольку эти животные питаются сухой пищей, содержащей крайне мало свободной воды, почти единственным источником воды для них является так называемая «метаболическая вода», которая получается при окислении питательных веществ. Этим очень небольшим количеством воды они должны возмещать все потери влаги с выдыхаемым воздухом, мочой и калом. Потери с мочой и калом сведены к минимуму благодаря тому, что животное выделяет очень концентрированную мочу и очень сухие фекалии, однако возможности уменьшить потери воды с выдыхаемым воздухом более ограничены, так как слизистые дыхательных путей всегда влажные и выдыхаемый воздух насыщен водяными парами. Таким образом, испарение — главный и притом неизбежный путь потери воды у этих животных. У человека, находящегося в пустыне, испарение гораздо выше, поскольку в жарких условиях потоотделение усиливается, что способствует охлаждению тела. Если бы грызуны, живущие в пустыне, потели, то все запасы воды в их организме {10} быстро иссякли бы; но потовых желез у них нет, и поскольку они ведут ночной образ жизни, проводя жаркое дневное время в норах, где относительно прохладно, они вообще обходятся без потоотделения.
Обычно считают, что выдыхаемый воздух насыщен водой, температура которой близка к температуре тела, составляющей для млекопитающих и птиц в среднем 37°С. С пресмыкающимися дело обстоит иначе; они относятся к так называемым «холоднокровным животным», а это значит, что температура их тела изменяется в соответствии с изменениями температуры среды. Температура тела (а следовательно, и количество воды, содержащейся в выдыхаемом воздухе) колеблется у них гораздо сильнее, а кроме того, она обычно гораздо ниже, чем у птиц и млекопитающих.
Для того чтобы сравнивать испарение воды у разных животных, удобно иметь какую-нибудь подходящую единую меру, ибо животные резко различаются по своим размерам и по потребностям в кислороде (а тем самым и по интенсивности легочной вентиляции). Поскольку легочная вентиляция, а следовательно и испарение, изменяется в зависимости от потребности в кислороде, такой удобной мерой может служить количество испаряемой воды на единицу объема потребляемого кислорода. Эта мера представляется особенно полезной, если вспомнить, что количество поглощенного кислорода служит непосредственным показателем количества воды, образующейся при метаболизме, которая составляет практически всю воду, поступающую в организм пустынных грызунов.
Если сравнить испарение воды разными животными (табл. 2), то окажется, что у тех видов, которые обитают в пустыне, оно гораздо ниже, чем у животных, живущих в других местообитаниях, или у человека. В чем причина такой разницы?
Принято считать, что температура тела и легочная вентиляция у разных видов примерно одинаковы. Как же в таком случае возникают различия в испарении? Следует отметить, что данные таблицы 2 относятся
| {11} |
Таблица 2
Испарение у разных видов
Для получения сравнимых данных количество испаряемой воды
отнесено к количеству поглощаемого за это же время
кислорода. Для грызунов указано суммарное количество
влаги, выделяемой с выдыхаемым воздухом и через кожу,
а для человека — только то количество, которое
выделяется через легкие (по данным Шмидта-Нильсена, 1964).
Вид |
Испарение, мг H2O |
Кенгуровая крыса |
0,54 ±0,04 |
Гофер |
0,50 ±0,07 |
Золотистый хомячок |
0,59 ±0,05 |
Белая крыса |
0,94 ±0,09 |
Белая мышь |
0 85 ±0,07 |
Человек |
0,84 |
к потерям воды не только с выдыхаемым воздухом, но и с поверхности тела (за исключением данных для человека, у которого испарение влаги с поверхности кожи довольно велико и сильно колеблется). Неоднократно высказывалось мнение, что у грызунов испарение влаги с поверхности тела очень незначительно, так как у них нет потовых желез; на самом же деле у белых крыс через кожу выделяется довольно большое количество влаги — около половины общего испарения (Теннент, 1946). Если исключить это количество, то испарение у белой крысы окажется примерно равным испарению у других мелких грызунов, то есть различие между пустынными и другими видами будет устранено.
У человека испарение, происходящее при дыхании, определяется насыщением воздуха влагой при температуре, близкой к температуре тела. Если это принять за эталон, то интересующий нас вопрос несколько видоизменяется. Каким образом животным удается уменьшить это испарение? Здесь возможны два способа: 1) из находящегося в легких воздуха, прежде чем животное его выдыхает, извлекается большее, чем {12} обычно, количество кислорода; 2) выдыхаемый воздух не насыщен влагой при температуре тела.
Первый способ кажется довольно приемлемым, во всяком случае более вероятным, чем второй. В легких из воздуха можно извлекать больше кислорода при условии повышения его сродства к содержащемуся в крови гемоглобину. Именно так обстоит дело у некоторых животных: например, у гуанако гемоглобин обладает высоким сродством к кислороду, что облегчает этому животному получение кислорода в разреженной атмосфере его высокогорных местообитаний в Андах (Холл, Дилл и Гузман Баррон, 1936). При высоком сродстве к гемоглобину кривая диссоциации кислорода сдвигается влево, однако для кенгуровой крысы такого сдвига не обнаружено; напротив, кривая диссоциации располагается у нее даже несколько правее, что свидетельствует о низком сродстве к кислороду, обычном для мелких млекопитающих.
Одного лишь этого факта недостаточно, чтобы отвергнуть возможность повышенного извлечения кислорода из воздуха в легких, но есть и другие данные, заставляющие отказаться от нее. Если бы из находящегося в легких воздуха извлекалось вдвое больше кислорода, чем обычно, то содержание углекислоты в выдыхаемом воздухе также стало бы вдвое выше. При этом напряжение углекислоты в крови должно было бы удвоиться, что привело бы либо к резко выраженному ацидозу, либо к изменению карбонатной буферной системы. Между тем буферная система крови у кенгуровой крысы ничем не отличается от аналогичной системы у других млекопитающих, и поэтому мы вынуждены отказаться от гипотезы повышенного извлечения кислорода (Ионнес и Шмидт-Нильсен, 1952).
Рассмотрим теперь другую возможность: выдыхаемый воздух не насыщен влагой при температуре тела. Изучая этот вопрос, мы обнаружили, что, хотя воздух насыщен влагой, его температура гораздо ниже температуры тела. В легких температура воздуха соответствует температуре тела, однако на пути из легких наружу воздух значительно охлаждается, что сопровождается конденсацией водяных паров на стенках дыхательных путей.
| {13} |
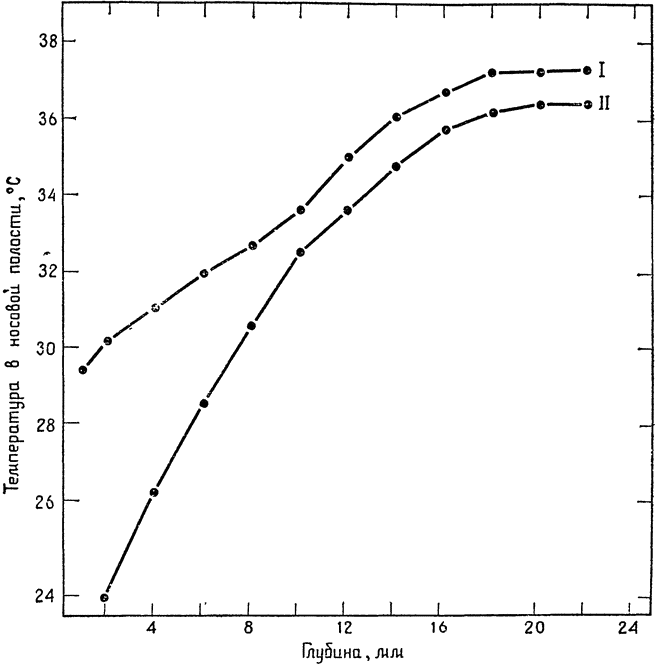
Измеряя температуру воздуха в носовой полости крысы, мы обнаружим, что она ниже всего у кончика носа, а по мере удаления от поверхности постепенно
 |
|
Рис. 1. Температура воздуха в носовой полости кенгуровой крысы (Dipodomys merriami) (по Джексону и Шмидту-Нильсену, 1964). I — при 100%-ной относительной влажности вдыхаемого воздуха и температуре 28°С; II — при вдыхании совершенно сухого воздуха и температуре 28°С. Температура тела животного 38°С. |
повышается, пока на глубине 20 мм не достигнет температуры тела (рис. 1). Когда животное вдыхает воздух, насыщенный влагой, температура у кончика носа приближается к температуре среды; однако в сухой атмосфере она на несколько градусов ниже температуры воздушной среды. Воздух, проходящий через {14} носовую полость, достигает (с отклонениями в пределах нескольких десятых градуса) той же температуры, что и у стенок этих полостей; в результате в сухой атмосфере животное выдыхает воздух, имеющий даже белее низкую температуру, чем тот, который оно вдыхает. Следует, конечно, помнить, что на выходе из легких воздух имеет температуру, равную температуре тела, то есть 38°С, и охлаждается при прохождении через носовую полость.
Для того чтобы понять, каким образом устанавливается температурный градиент в носовой полости,
 |
|
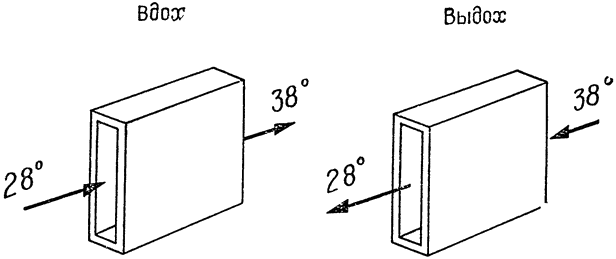
Рис. 2. Модель теплообмена, происходящего в носовой полости (по Шмидту-Нильсену и др., 1970, б). Окружающий воздух насыщен влагой и имеет температуру 28°С; температура тела равна 38°С. Вдыхаемый воздух, проходя через носовую полость, поглощает тепло и водяные пары и к тому времени, когда он попадает в легкие, насыщен влагой и нагрет до 38°С. При выдыхании воздух протекает по более холодным стенкам полости, отдавая тепло и влагу. При завершении теплообмена и водного обмена температура выдыхаемого воздуха приближается к 28°С и содержание в нем влаги соответствует точке насыщения для данной температуры. |
рассмотрим простую модель носа (рис. 2). Чтобы не усложнять дела, допустим, что испарения не происходит. При вдохе стенки носовой полости охлаждаются проходящим по ним воздухом и их температура приближается к температуре вдыхаемого воздуха, в данном случае — к 28°С. При выдохе теплый воздух из легких, проходя над этими поверхностями, охлаждается, и, при условии полного теплообмена, температура выдыхаемого воздуха приблизится к температуре стенок носовой полости, то есть к 28°С. Измерения, проведенные на кенгуровой крысе во влажной атмосфере (рис. 1, кривая I), дали примерно такую картину — температура выдыхаемого воздуха {15} приближалась к температуре среды. В совершенно сухой атмосфере (также при 28°С) температура выдыхаемого воздуха составляла около 23°С, то есть была на 5° ниже температуры среды (рис. 1, кривая II). Эта разница объясняется тем, что при вдыхании из влажной слизистой носа испаряется вода, в результате чего температура слизистой становится ниже температуры воздуха — подобно тому, как это происходит с влажным термометром в психрометре.
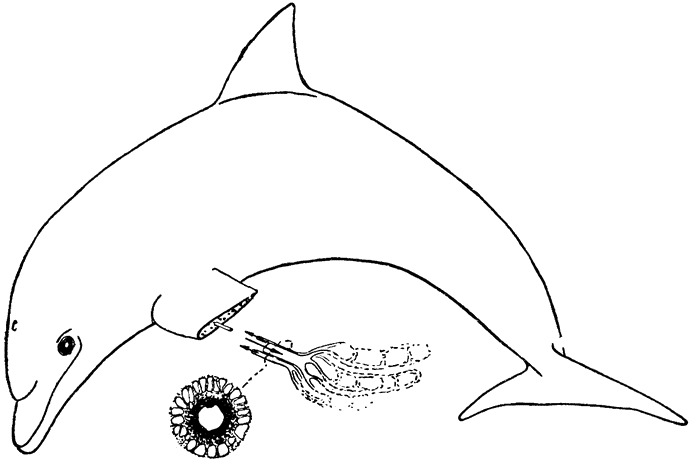
Теплообмен, происходящий в носовой полости, очень сходен с хорошо известным противоточным теплообменом, происходящим в конечностях многих водных животных, например в плавниках китов и ногах болотных птиц. Тело кита, плавающего в воде, температура которой близка к точке замерзания, покрыто толстым слоем подкожного жира, обеспечивающим теплоизоляцию. Но тонкие обтекаемой формы плавники и ласты не имеют такого теплоизолирующего слоя, а кроме того, они обильно снабжены кровеносными сосудами; поэтому животное теряло бы через плавники очень много тепла, если бы не теплообмен между артериальной и венозной кровью, происходящий в этих конечностях. Сосуды, по которым холодная венозная кровь возвращается из плавника к телу, проходят очень близко от артерий, практически окружая последние; благодаря такому расположению артериальная кровь отдает тепло венозной крови, которая таким образом нагревается, прежде чем поступить в тело (рис. 3). В конечностях многих других животных артерии и вены разветвляются на множество параллельных переплетающихся между собой сосудов диаметром около 1 мм каждый, образуя густую сеть (Шоландер, 1958; Шоландер и Крог, 1957). Образуют ли кровеносные сосуды такую сеть или просто располагаются в тесной близости друг к другу, принципиального значения не имеет, так как самый механизм теплообмена во всех случаях одинаков: по артериям и венам кровь течет в противоположных направлениях и между этими двумя параллельными системами {16} трубок происходит обмен тепла; поэтому механизм в целом известен под названием противоточного теплообменника.
Теплообмен в носовой полости основан на аналогичном принципе. В конечности кровь течет в противоположных направлениях по двум обособленным трубкам, а в носу вдыхаемый и выдыхаемый воздух проходит по одной и той же полости. Однако в носу
 |
|
Рис. 3. Потери тепла из конечностей китообразных снижаются благодаря особому расположению кровеносных сосудов. На поперечном сечении плавника видно, что артерия со всех сторон окружена венами. В результате теплая артериальная кровь отдает тепло венозной крови, которая таким образом нагревается, прежде чем вновь поступить из плавников в тело. |
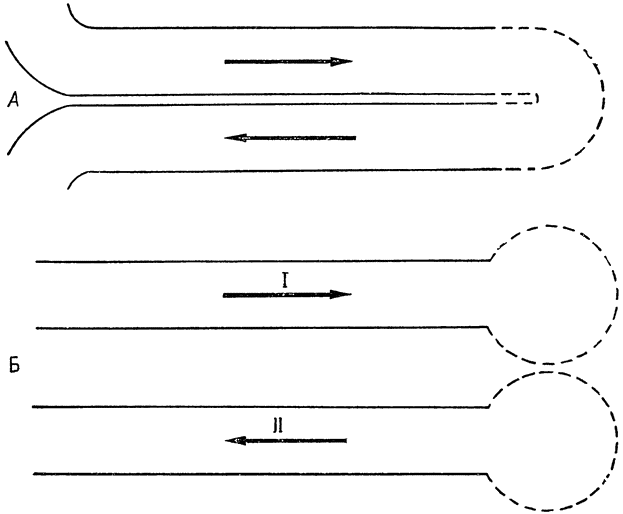
эти два потока разделены во времени, так что мы можем говорить о временном разделении противоточного теплообменника, тогда как в конечностях кровеносные сосуды разделены пространственно. Это различие поясняют схемы на рис. 4.
Рассмотрим теперь общие принципы обмена тепла в дыхательных путях и его значение для поддержания водного баланса у млекопитающих и птиц. Подобная система теплообмена свойственна не только животным пустыни; эта система — неизбежное следствие геометрии дыхательных путей, и мы попытаемся {17} проанализировать ее здесь в общих чертах. Мы намерены также показать, почему даже холоднокровным животным выгодно охлаждение выдыхаемого воздуха, и, наконец, предложить использовать принцип
 |
|
Рис. 4. Модели противоточных теплообменников (по Шмидту-Нильсену и др., 1970, б). В плавниках кита или в ногах птиц кровь течет по двум трубкам, разделенным в пространстве (пространственное разделение). Б. В носовой полости при вдыхании (I) ее стенки отдают тепло, а при выдыхании (II) воздух, протекающий в обратном направлении, возвращает им тепло. Таким образом, теплообменник, находящийся в носу, состоит из одной трубки, а два потока воздуха разделены во времени (временное разделение). |
функционирования теплообменника в носу кенгуровой крысы для создания тепловой машины, которая представляет собой реализацию принципа вечного двигателя.
У птиц температура выдыхаемого воздуха также несколько ниже температуры тела и приближается к температуре среды. Мы провели соответствующие измерения на кактусовых вьюрках (которые по размерам близки к кенгуровым крысам и весят примерно {18} 35 г) в диапазоне температур окружающей среды от 12 до 30°С (рис. 5). Выдыхаемый воздух всегда имел несколько более низкую температуру, чем сама птица (41°С), и разница между температурой воздуха и температурой тела колебалась от 8 до 23°С (Шмидт-Нильсен,
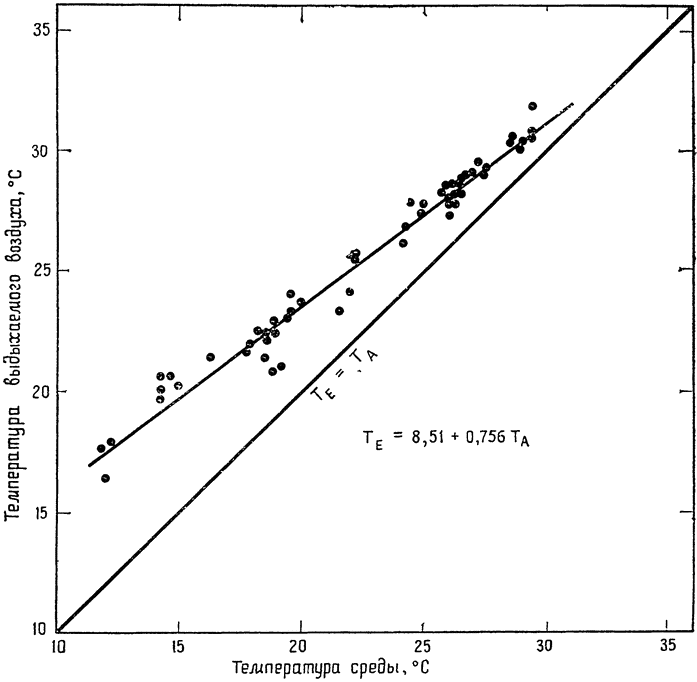 |
Рис. 5. Температура воздуха, выдыхаемого кактусовым вьюрком (35 г), при колебаниях температуры среды от 12 до 30°С; температура тела вьюрка все время составляла 41°С (по Шмидту-Нильсену и др., 1970, б). |
Эйнсуорт и Марриш, 1970,6). Иными словами, по всему диапазону колебаний температуры окружающей среды разность температур между вдыхаемым и выдыхаемым воздухом не выходила за пределы нескольких градусов (1—7°С). Кактусовый вьюрок, подобно кенгуровой крысе, — обитатель пустынь; охлаждение выдыхаемого воздуха снижает {19} испарение влаги через дыхательные пути, что, несомненно, благоприятно сказывается на водном балансе этих птиц.
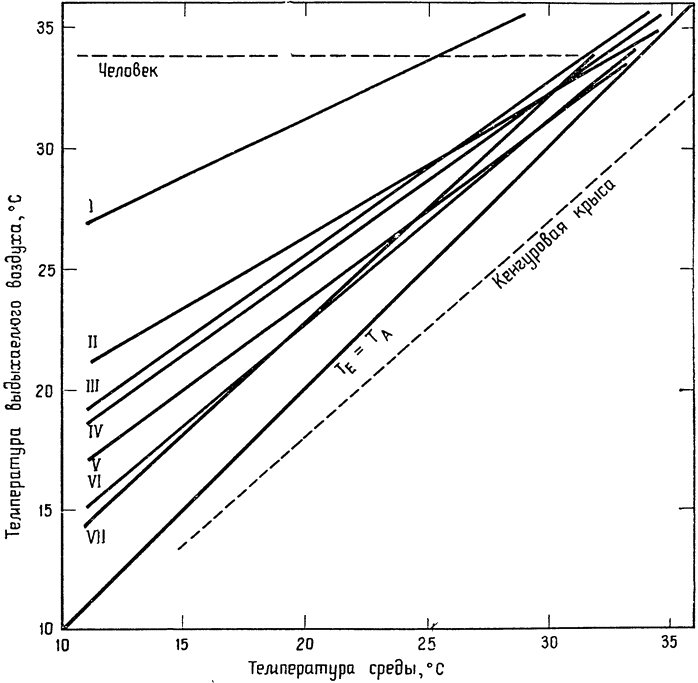 |
|
Рис. 6. Температура выдыхаемого воздуха (ТЕ) в зависимости от температуры среды (ТА) у семи видов птиц (по Шмидту-Нильсену и др., 1970, б). I — домашняя утка; II — домовый воробей; III — австралийский волнистый попугайчик; IV — голубь; V — кактусовый вьюрок; VI — каролинский вьюрок; VII — дрозд-отшельник. Линии регрессии построены по данным, аналогичным приведенным на рис. 5, и вычислены по методу наименьших квадратов. Линия для кенгуровой крысы показывает, что охлаждение выдыхаемого воздуха происходит у нее более эффективно, чем у птиц. Линия для человека вычерчена по данным, полученным на одном индивидууме; измерения, проведенные на других индивидуумах, дали бы несколько иные линии, обычно с положительным наклоном. |
Следовало ожидать, что, сравнивая птиц, обитающих в пустыне, с птицами, живущими в других местах, мы обнаружим между ними какие-то различия, способствующие сохранению воды у пустынных видов. {20} Однако результаты измерений по семи видам птиц не дают оснований считать, что птицы пустыни в чем-то отличаются от обитателей других мест (рис. 6). Оказалось, что у кактусового вьюрка температура выдыхаемого воздуха даже несколько выше, чем у его близкого родича — Каролинского вьюрка. Кроме того, австралийский волнистый попугайчик (типичный обитатель засушливых мест) практически не отличался в этом отношении от нашего обычного домового воробья.
Один вид птиц — домашняя утка — резко отличался от всех остальных: температура выдыхаемого воздуха у нее была значительно выше, чем у других видов. Это, несомненно, объясняется тем, что у уток носовая полость гораздо шире, чем у других видов, и поэтому условия для теплообмена менее благоприятны. У человека температура выдыхаемого воздуха также значительно выше (хотя температура тела у него на 3—4° ниже, чем у птиц), и обычно составляет 34—35°С. Причина та же: в широкой носовой полости теплообмен между выдыхаемым воздухом и ее поверхностями затруднён.
У кенгуровой крысы, как говорилось выше, выдыхаемый воздух охлаждается до температур ниже температуры среды, и ее линия регрессии располагается поэтому гораздо ниже группы линий для птиц, близких к ней по размерам. Это различие между птицами и кенгуровой крысой легко понять, если сравнивать строение носовой полости и условия для теплообмена между выдыхаемым воздухом и стенками этой полости у птиц и у крысы. На поперечном срезе носа кенгуровой крысы (рис. 7) можно видеть, что сама полость представляет собой довольно узкий проход, а ее стенки занимают много места; подобное строение создает оптимальные условия для обмена тепла между проходящим через носовую полость воздухом и окружающими тканями. У птиц полость шире и короче, площадь поверхности меньше и условия для теплообмена менее благоприятны. Особенно широка носовая полость у уток. У человека же теплообмен между воздухом и стенками носовой полости настолько незначителен, что в большинстве случаев он почти не играет никакой роли. Таким образом, положение о том, что содержание влаги в выдыхаемом воздухе {21} достигает точки насыщения при температуре тела или очень близко к ней, совершенно справедливо для человека и соответствует наблюдаемым фактам. Ошибка
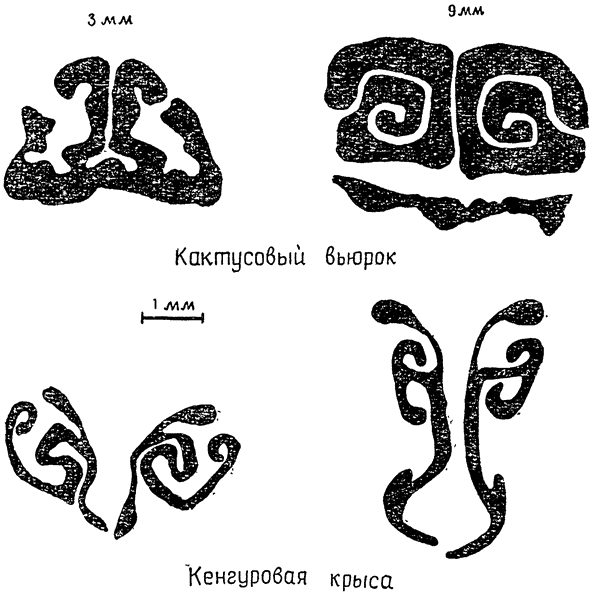 |
|
Рис. 7: Поперечные срезы-через носовую полость кактусового вьюрка и кенгуровой крысы (по Шмидту-Нильсену и др., 1970, б). У птицы полости (черные) шире, а стенки (белые) тоньше, чем у млекопитающего того же размера (вес тела около 35 г). Срезы сделаны на глубине 3 и 9 мм от наружных ноздрей. |
состояла в обобщении этого положения применительно ко всем теплокровным животным.
Прежде чем рассматривать значение охлаждения выдыхаемого воздуха для уменьшения потерь воды организмом, полезно напомнить, что содержание водяных паров в насыщенном влагой воздухе сильно зависит от температуры. Как показывает кривая на рис. 8, количество водяных паров, которое может содержать воздух, с повышением температуры быстро {22} возрастает. Можно указать одно легко запоминающееся эмпирическое правило: насыщенный влагой воздух при температуре тела содержит ровно вдвое больше водяных паров (46 мг/л), чем насыщенный воздух при комнатной температуре (23 мг/л). Таким образом, при охлаждении выдыхаемого воздуха от температуры тела до комнатной температуры количество водяных паров, которые он может унести с собой,
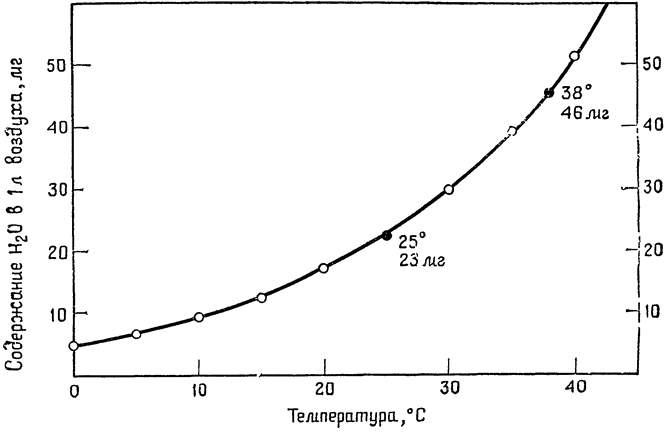 |
|
Рис. 8. Содержание водяных паров в воздухе при полном насыщении (100%-ная относительная влажность). Наклон кривой становится все более крутым по мере приближения к температуре, соответствующей температуре тела птиц и млекопитающих, то есть количество влаги, необходимое для насыщения находящегося в легких воздуха, сильно возрастает. |
уменьшается вдвое — довольно существенная экономия воды, если вспомнить, что у животных, о которых идет речь, организм теряет воду главным образом за счет испарения.
Влияние охлаждения выдыхаемого воздуха на водный баланс кенгуровой крысы показано на рис. 9 (диаграмма в принципе пригодна и для других мелких грызунов). Расчеты сделаны для двух произвольно выбранных температур окружающего воздуха — 15 и 30°С; относительная влажность в обоих случаях произвольно принята равной 25%. Заштрихованные столбцы показывают количество водяных паров, {23} которое должно быть добавлено к 1 л наружного воздуха после его вдыхания, чтобы содержание в нем влаги достигло точки насыщения при температуре, равной температуре тела (38°С). Незаштрихованные столбцы расположены над точками, соответствующими температуре выдыхаемого воздуха при выбранных условиях. Если, например, температура вдыхаемого
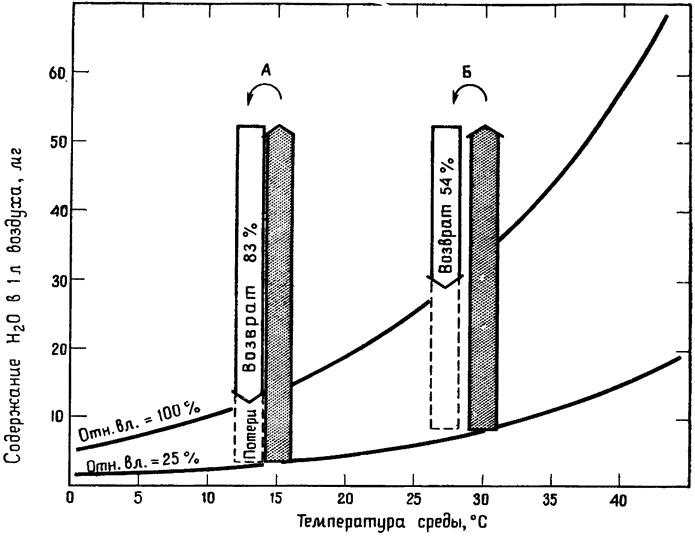 |
Рис. 9. Диаграмма процесса извлечения воды из выдыхаемого воздуха у кенгуровой крысы (по Шмидту-Нильсену и др., 1970,6). Объяснения в тексте. |
воздуха равна 15°С, то температура выдыхаемого воздуха составит 13°С (рис. 9, A). В этом случае охлаждение выдыхаемого воздуха позволяет удержать в организме 83% влаги, добавленной к воздуху при его вдыхании, то есть животное теряет лишь около 1/6 воды, необходимой для насыщения вдыхаемого воздуха. При 30°С удается сэкономить меньше воды, но даже в этом случае в организме удерживается 54% той воды, которая была бы потеряна, если бы температура выдыхаемого воздуха была равна температуре тела. {24}
У птиц, типичным примером которых может служить кактусовый вьюрок, экономия воды будет меньше, так как выдыхаемый воздух охлаждается менее эффективно, но все же эта экономия довольно существенна. В условиях, сходных с теми, в которых производились измерения на кенгуровой крысе (15°С и 25%-ная относительная влажность), вьюрку удается сохранить 74% воды, а при температуре 30°С — 49% (Шмидт-Нильсен и др., 1970, б). Конечно, следует помнить, что температура тела, у птиц выше, чем у млекопитающих (41 и 38°С), и для насыщения находящегося в их легких воздуха нужно больше влаги.
Какое влияние оказывает сэкономленная вода на тепловой бюджет этих мелких животных? В таблице 3 показано, как распределяется тепло, которое кенгуровая крыса отдает воздуху при вдыхании, и какую его часть она вновь получает при выдыхании. Оказывается, при 15°С и 25%-ной относительной влажности на нагревание и увлажнение 1 л вдыхаемого воздуха расходуется 35 кал тепла, из которых 31 кал возвращается организму при выдыхании. Таким образом, при этих условиях около 88% тепла, используемого на нагревание и увлажнение выдыхаемого воздуха, организм получает назад, то есть тепло, затрачиваемое на эти процессы, не утекает из организма, что весьма существенно, поскольку это составляет 16% тепла,
Таблица 3
Сравнение количества тепла, расходуемого кенгуровой крысой
на нагревание и увлажнение 1 л вдыхаемого воздуха
(температура 15°С, относительная влажность 25%),
с количеством тепла, получаемым организмом за счет
охлаждения этого же самого воздуха при выдыхании
Расход при |
Возврат при |
Экономия при |
|
Теплота испарения |
28,5 |
23,5 |
83 |
Нагревание воздуха |
6,8 |
7,4 |
107 |
Суммарное тепло |
35,3 |
30,9 |
88 |
| {25} |
образуемого организмом за счет метаболизма при вдыхании 1 л воздуха (192 кал). При более высоких температурах среды относительное значение экономии тепла будет меньше, но при низких температурах его роль возрастает.
У человека теплообмен между потоком вдыхаемого воздуха и стенками носовой полости гораздо менее эффективен, поскольку полость эта гораздо шире, а поток воздуха имеет ламинарный характер и расстояние от ядра потока до стенок полости слишком велико. Температура выдыхаемого человеком воздуха близка к температуре его тела, а возврат тепла и водяных паров обычно незначителен. Однако при очень низких температурах потери тепла, связанные с нагреванием и увлажнением вдыхаемого воздуха, становятся существенной «статьей расхода» даже в тепловом бюджете человека. Если, например, человек дышит сухим воздухом при температуре 0°С, то примерно 20% тепла, образуемого им за счет метаболизма, расходуется на нагревание и увлажнение вдыхаемого воздуха. При еще более низких температурах воздуха дыхательные пути начинают сильно охлаждаться; так, при —30°С извлечение тепла и воды из выдыхаемого воздуха становится весьма важным фактором в уменьшении утечки тепла из организма (Уокер, Уэллс и Меррилл, 1961; Уэбб, 1951, 1955).
Пытаясь изучать носовую полость как теплообменник, мы сталкиваемся с необходимостью учитывать огромное число переменных. При переносе тепла между воздухом и стенками полости вода либо испаряется, либо конденсируется, и при этом происходит либо связывание, либо освобождение значительных количеств скрытой теплоты парообразования. Для потоков тепла и воздуха совершенно очевидна важность таких геометрических переменных, как общая поверхность, на которой может происходить теплообмен, и расстояние от ядра потока воздуха до стенок полости. Важную роль играют и такие физические условия среды, как температура воздуха и влажность, {26} поскольку оба эти условия оказывают влияние на перенос тепла и испарение. Необходимо учитывать также ряд физиологических переменных. Интенсивность дыхания и объем вдыхаемого воздуха могут варьировать; оба эти параметра воздействуют на линейную скорость протекающего через носовую полость воздушного потока, а скорость потока непрерывно изменяется на протяжении каждого цикла дыхания. Наконец, не следует забывать и о кровотоке, который не только поставляет испаряющуюся воду, но и переносит тепло от внутренних участков тела к области носа.
При вдыхании тепло распространяется в поперечном направлении — от стенок носовой полости к воздушному потоку — и водяные пары поступают в воздух по создающемуся таким образом поперечному температурному градиенту. При выдыхании происходит обратный процесс: тепло передается от воздуха стенкам носовой полости, и по мере снижения температуры водяные пары рассеиваются в поперечном направлении, вновь конденсируясь на этих стенках. Адекватное описание этих процессов при непрерывно изменяющихся линейных скоростях потоков на протяжении одного цикла дыхания оказалось совершенно невыполнимой задачей.
Один из моих бывших учеников, Дж. Коллинз, показал, что всем этим нескончаемым числом переменных можно пренебречь и представить носовую полость в виде относительно простой стационарной модели. На рис. 10 схематически изображена такая модель; штриховой линией обозначены воображаемые границы носовой полости и ее влажной выстилки, начинающиеся на некотором расстоянии от кончика носа (граница х) и тянущиеся по границе у до границы z, где температура равна температуре глубоких областей тела. По этой полости проходит также поток жидкости (крови). Энергию, переносимую через указанную границу, можно мысленно разделить на несколько составных частей. Прежде всего следует указать, что содержание энергии в воздухе, пересекающем границу z, не изменяется, поскольку эта граница по определению расположена там, где температура воздуха равна температуре тела и он насыщен влагой. Поток энергии, который проходит через границы х и у, слагается из {27} четырех компонентов:
a) изменение температуры воздуха;
b) испарение воды;
c) теплоотдача во внешнюю среду;
d) изменение температуры крови.
Это позволяет построить математическую модель, описывающую устойчивое энергетическое равновесие системы. В этой модели энергия, переносимая из потока жидкости (d — кровоток и перенос тепла от глубоких областей тела к области носа), равна суммарной
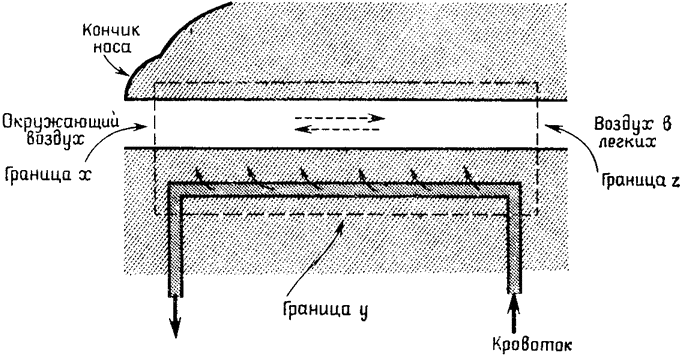 |
Рис. 10. Схема стационарной модели, позволяющей изучать обмен воды и тепла в носовой полости. Объяснения в тексте. |
энергии, переносимой за счет: а — изменения температуры вдыхаемого воздуха, b — испарения и с — непосредственной отдачи тепла во внешнюю среду, происходящей на самом кончике носа, то есть а + b + с = d. Все переменные, входящие в это уравнение, за исключением d, могут быть измерены или оценены на основании физиологических данных, что позволяет решить его относительно скорости потока жидкости, Получив эмпирически значение для скорости потока жидкости, можно исключить эту переменную и вывести общее уравнение энергетического равновесия, которое в свою очередь позволяет вычислить значения температуры выдыхаемого воздуха, скорости потери воды и эффективности сохранения водяных паров для различных внешних условий (температуры и влажности воздушной среды). {28}
Я не буду здесь касаться проверки описанной математической модели с помощью подлинных физиологических измерений (это было подробно описано в работе Коллинза, Пилкингтона и Шмидта-Нильсена, 1971), а ограничусь тем, что приведу две таблицы, в которых содержатся предсказанные моделью температуры носа кенгуровой крысы при разных атмосферных условиях и доля водяных паров, возвращаемых организму благодаря конденсации на стенках носовой полости. В данных таблицы 4, где приведены температуры
Таблица 4
Предсказанная математической моделью зависимость
температуры носа кенгуровой крысы от условий среды
(Коллинз и др., 1971)
Темпера- |
Относительная влажность среды, % |
|||||
0 |
20 |
40 |
60 |
80 |
100 |
|
10 |
9,8 |
11,8 |
13,7 |
15,3 |
16,8 |
18,1 |
12 |
10,7 |
12,9 |
14,8 |
16,5 |
18,1 |
19,5 |
14 |
11,7 |
14,0 |
16,0 |
17,8 |
19,4 |
20,9 |
16 |
12,6 |
15,1 |
17,2 |
19,1 |
20,8 |
22,4 |
18 |
13,6 |
16,2 |
18,4 |
20,4 |
22,2 |
23,9 |
20 |
14,6 |
17,3 |
19,7 |
21,7 |
23,6 |
25,3 |
22 |
15,6 |
18,5 |
21,0 |
23,1 |
25,1 |
26,9 |
24 |
16,6 |
19,6 |
22,2 |
24,5 |
26,5 |
28,4 |
26 |
17,7 |
20,8 |
23,5 |
25,9 |
28,0 |
29,9 |
28 |
18,8 |
22,0 |
24,9 |
27,3 |
29,5 |
31,4 |
30 |
19,7 |
23,2 |
26,1 |
28,6 |
30,9 |
33,0 |
32 |
20,6 |
24,3 |
27,4 |
30,0 |
32,4 |
34,5 |
34 |
21,6 |
25,4 |
28,6 |
31,4 |
33,8 |
36,0 |
36 |
22,6 |
26,6 |
29,9 |
32,8 |
35,3 |
37,6 |
носа, предсказанные моделью, нет ничего неожиданного: температура выдыхаемого воздуха в значительной степени зависит от условий среды, причем особенно важное значение имеет относительная влажность вдыхаемого воздуха. При высокой влажности выдыхаемый воздух, как и следовало ожидать, в среднем охлаждается не так сильно, как в сухой атмосфере. {29}
Более неожиданными оказались предсказанные моделью данные о возврате воды, происходящем в носовой полости (табл. 5). В широком диапазоне
Таблица 5
Предсказанная математической моделью зависимость эффективности возвращения воды в организм кенгуровой крысы от условий среды (Коллинз и др., 1971)
Темпера- |
Относительная влажность среды, % |
|||||
0 |
20 |
40 |
60 |
80 |
100 |
|
10 |
0,755 |
0,756 |
0,758 |
0,760 |
0,763 |
0,766 |
12 |
0,749 |
0,750 |
0,753 |
0,755 |
0,758 |
0,762 |
14 |
0,743 |
0,745 |
0,747 |
0,750 |
0,754 |
0,757 |
16 |
0,736 |
0,739 |
0,741 |
0,745 |
0,749 |
0,753 |
18 |
0,730 |
0,732 |
0,735 |
0,739 |
0,743 |
0,748 |
20 |
0,723 |
0,725 |
0,729 |
0,733 |
0,737 |
0,743 |
22 |
0,715 |
0,718 |
0,722 |
0,726 |
0,731 |
0,737 |
24 |
0,707 |
0,710 |
0,714 |
0,719 |
0,725 |
0,731 |
26 |
0,698 |
0,702 |
0,706 |
0,711 |
0,717 |
0,724 |
28 |
0,688 |
0,692 |
0,697 |
0,703 |
0,709 |
0,715 |
30 |
0,682 |
0,685 |
0,692 |
0,697 |
0,703 |
0,710 |
32 |
0,675 |
0,680 |
0,685 |
0,691 |
0,697 |
0,704 |
34 |
0,667 |
0,672 |
0,678 |
0,684 |
0,690 |
0,696 |
36 |
0,659 |
0,665 |
0,671 |
0,677 |
0,682 |
0,685 |
температур воздуха (от 10 до 36°С) и относительной влажности (от 0 до 100%) среды предсказанная доля возвращаемой организму воды колебалась лишь от 66 до 77%. Иными словами, модель предсказывает, что, несмотря на значительные колебания условий среды, доля возвращаемой организму воды будет оставаться примерно постоянной, составляя около 75%.
Для того чтобы установить этот факт без помощи вычислительной машины, пришлось бы провести огромное количество однообразных и довольно скучных лабораторных наблюдений. Хотя возможности модели для предсказания результатов несколько ограничены, {30} нам кажется целесообразным поручить эту работу вычислительной машине, а самим заняться другими проблемами.
Измерив температуру воздуха, выдыхаемого ящерицей, мы убедимся, что она очень близка к температуре ее тела (рис. 11). Это неудивительно, так как
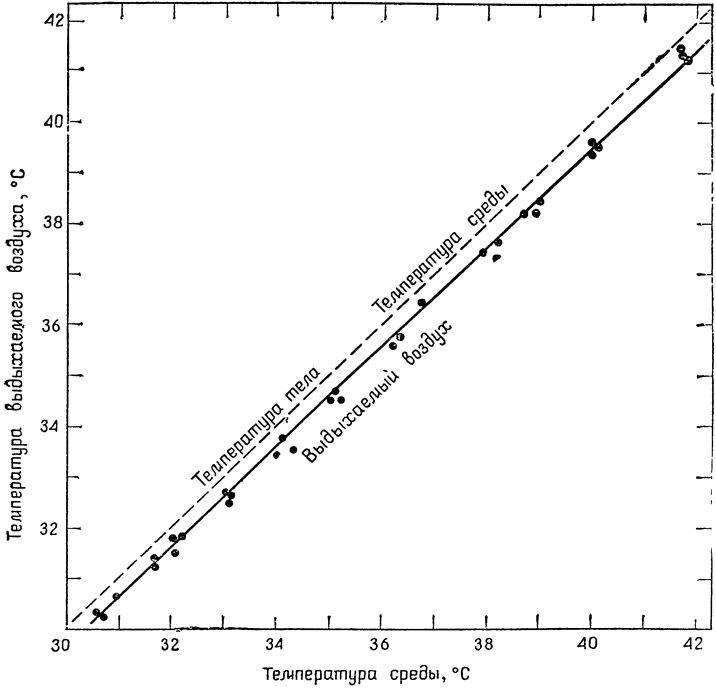 |
Рис. 11. Температура воздуха, выдыхаемого пустынной игуаной (Dipsosaurus dorsalis) (по Марришу и Шмидту-Нильсену, 1970). |
температура тела ящерицы примерно соответствует температуре ее среды и наблюдаемое незначительное охлаждение выдыхаемого воздуха обусловлено влажностью внутренних поверхностей дыхательных путей {31} и некоторым их охлаждением в результате испарения. Пока что все происходит так, как и следовало ожидать, и испарение из дыхательных путей зависит, помимо их объема, только от температуры и влажности среды.
Осложнения возникают, однако, как только мы вспоминаем, что температура тела у ящериц далеко не всегда соответствует температуре окружающего воздуха; многие ящерицы способны повышать температуру своего тела до уровня, значительно превосходящего температуру среды, просто-напросто греясь на солнце. Поскольку вдыхаемый воздух насыщается водяными парами в легких, при высокой температуре тела для его насыщения потребуется больше воды. Когда температура тела отличается от температуры среды, температура воздуха, находящегося в легких, соответствует температуре глубоких областей тела, однако это не означает, что и выдыхаемый воздух имеет такую же температуру.
У пустынной игуаны (Dipsosaurus dorsalis) в естественных условиях температура тела нередко достигает 42°С, хотя температура окружающего воздуха может составлять лишь 30°С (Норрис, 1953). Оказывается, игуана не просто греется на солнце, но и принимает такие позы, при которых на ее тело падает максимальное количество солнечных лучей. В лабораторных условиях мы поддерживали температуру этих ящериц на уровне 42°С при помощи излучения, имитирующего солнечное, и измеряли температуру выдыхаемого ими воздуха при различных температурах среды. Оказалось, что выдыхаемый воздух охлаждается довольно значительно (рис. 12). Например, температура выдыхаемого воздуха была на 7°С ниже температуры тела, которая составляла 42°С, а температура воздуха в комнате была равна 30°С (Марриш и Шмидт-Нильсен, 1970). Именно в таких условиях могла бы оказаться игуана, греясь ранним утром на солнце в пустыне; наблюдавшееся охлаждение выдыхаемого воздуха снизило бы потери воды до 31% по сравнению с потерями, которые произошли бы в том случае, если бы температура выдыхаемого воздуха была такой же, как и в легких.
Охлаждение выдыхаемого воздуха у ящериц происходит по тому же принципу, что и у птиц и {32} млекопитающих, однако эффективность этого процесса у ящериц гораздо ниже и соответственно меньше экономия воды. Это объясняется прежде всего иной формой их носовой полости: она шире и короле, и общая
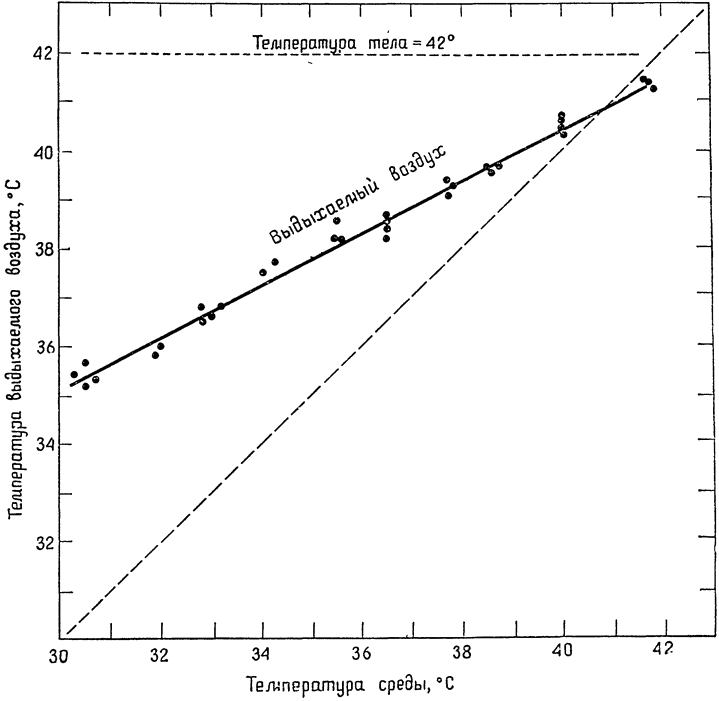 |
Рис. 12. Изменение степени охлаждения воздуха, выдыхаемого игуаной, у которой температура тела была равна 42°С, в зависимости от температуры окружающего воздуха (по Марришу и Шмидту-Нильсеку, 1970). |
площадь ее поверхности меньше, чем, например, у кенгуровой крысы.
Однако в строении носовой полости у пустынней игуаны есть одна особенность, которая влечет за собой некоторые весьма интересные последствия. Самая дистальная часть этой полости, расположенная сразу за наружными ноздрями, образует небольшую {33} впадину (рис. 13, 1); выделяемая носовыми солевыми железами жидкость накапливается во впадине и используется для увлажнения вдыхаемого воздуха. Па мере испарения воды эта жидкость становится все более концентрированной, из нее выпадают кристаллы соли, образующие инкрустации в отверстиях ноздрей или вокруг них. Таким образом, воду, содержащуюся
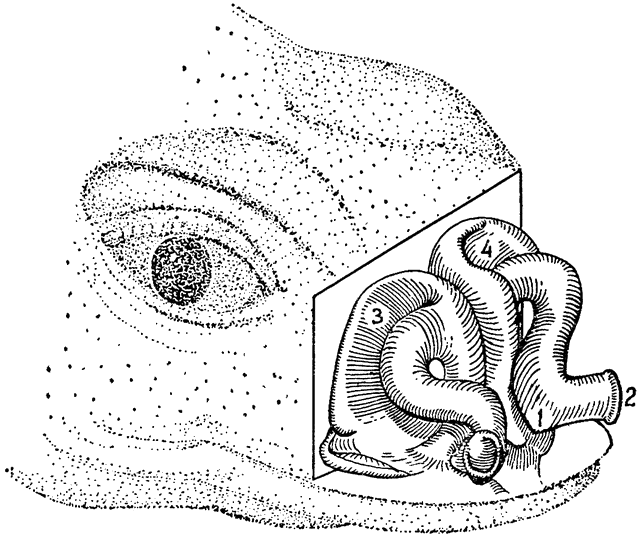 |
Рис. 13. В носовой полости пустынной игуаны имеется небольшая впадина (1), расположенная за наружными ноздрями (2). Жидкость из носовых солевых желез (3 и 4) накапливается в этой впадине и используется для увлажнения вдыхаемого воздуха (по Марришу и Шмидту-Нильсену, 1970). |
в секрете носовых желез, нельзя считать совершенно потерянной для организма. Поскольку так или иначе на увлажнение вдыхаемого воздуха должно быть затрачено какое-то количество воды, испарение из носовой полости дает возможность извлечь пользу из воды, которая уже, так сказать, «выведена» из организма. Иными словами, использование этой воды для увлажнения равносильно тому, что носовые железы секретировали бы соли в кристаллическом состоянии, без всякой воды,
| {34} |
На рис. 14 изображена схема машины, в которой используется примерно тот же принцип теплообмена, что и в носовой полости кенгуровой крысы. Машина состоит из двух воздухонепроницаемых цилиндров. Меньший цилиндр снабжен обыкновенным поршнем, а
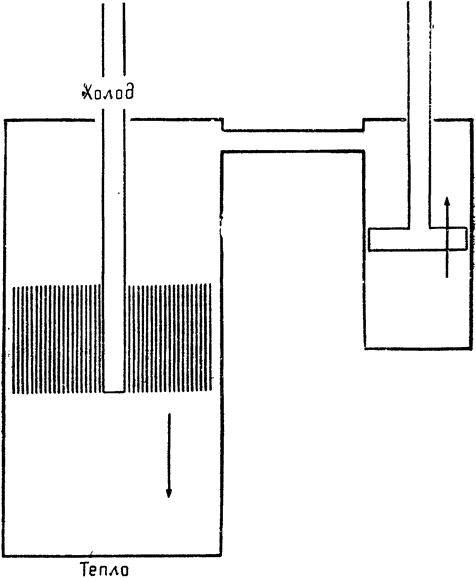 |
Рис. 14. Схема тепловой машины, которая на первый взгляд работает как вечный двигатель. |
в большем имеется поршень особого рода: он не прилегает к стенкам цилиндра вплотную, что позволяет уменьшить трение; кроме того, тело этого поршня состоит из вертикальных пластинок, между которыми может проходить воздух. Один конец большего цилиндра все время нагрет, а другой остается холодным. Если, как показано на рисунке, большой поршень опустить вниз (что не требует особых затрат энергии {35} вследствие неплотного прилегания поршня к стенкам цилиндра и свободного прохождения через него воздуха), то нагретый воздух в нижней части цилиндра отдает тепло пластинкам поршня. При достаточно эффективном теплообмене воздух, выходящий в верхнюю часть цилиндра, будет охлаждаться до температуры поршня. Когда большой поршень находится у основания, вся система заполнена холодным воздухом и снизившееся давление заставляет поршень в меньшем цилиндре двигаться вверх.
На следующей стадии цикла большой поршень движется в противоположном направлении; при этом холодный воздух, протекая между нагретыми пластинками поршня, отнимает у них тепло; поэтому, когда большой поршень находится в самом верхнем положении, больший цилиндр целиком заполнен нагретым воздухом. Возросшее давление толкает маленький поршень вниз, и цикл завершается. Поскольку при движении большого поршня трение незначительно и воздух свободно проходит между пластинками, для того чтобы поднимать и опускать этот поршень, не требуется больших затрат энергии. Эта энергия составляет некоторую долю энергии, создаваемой движением маленького поршня, а остальная часть последней может быть использована для выполнения той или иной работы.
Действие этой машины основано на теплообмене в большом поршне. Воздух получает тепло, а затем вновь отдает его при изменении направления потока в теплообменнике. На первый взгляд кажется, что такая машина представляет собой вечный двигатель. Потери на трение в большом поршне можно почти свести к нулю, так что эффективность теплообмена теоретически будет близка к 100%. Однако вслед за этим возникает мысль, что поскольку вечный двигатель создать невозможно, то, вероятно, вся эта схема основана на каком-то ошибочном допущении. Могу заверить вас, однако, что это не так. Такая тепловая машина была изобретена одним шотландским священником, преподобным Робертом Стерлингом, и запатентована им в 1827 году, а затем — уже в усовершенствованном виде — в 1840 году. Она была даже {36} построена и введена в действие (рис. 15)1. В технической литературе XIX века сообщается о тепловой
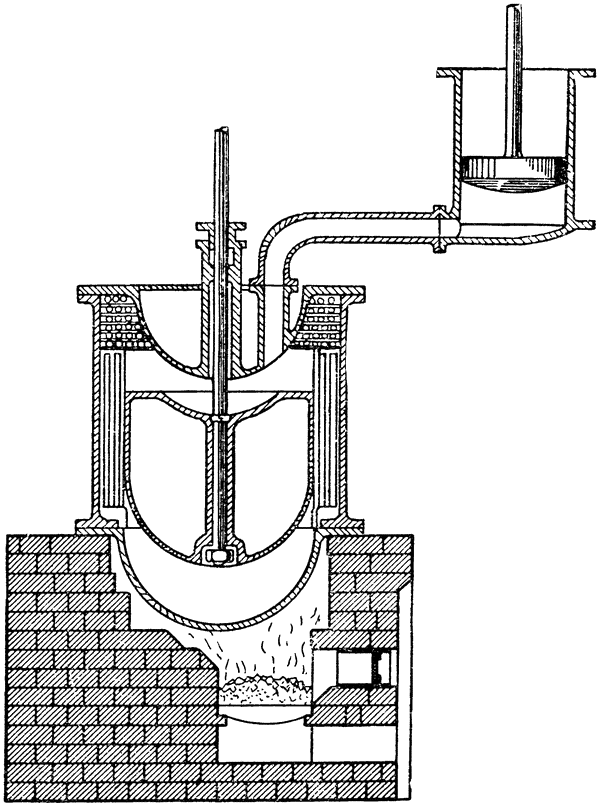 |
Рис. 15. Тепловая машина Стирлинга, работающая по принципу, показанному на рис. 14 (по Дженкину, 1885). |
машине Стирлинга мощностью в 50 лошадиных сил, которая на протяжении многих лет использовалась на {37} литейном заводе в Данди. Испытания термодинамической эффективности машины Стирлинга показали, что она подчиняется тем же законам, что и тепловая машина Карно; по существу оба цикла идентичны, отличаясь лишь тем, что машина Карно действует в адиабатических условиях, а машина Стирлинга — в условиях постоянного объема.
Мне хочется закончить эту главу указанием на то, что теплообменники или регенераторы тепла типа машины Стирлинга находят все более широкое применение; в частности, их используют для получения температур ниже температуры жидкого гелия (Джиффорд и Мак-Магон, 1960). Температуры, близкие к абсолютному нулю, легко получить при помощи теплообменников, построенных по тому же принципу, который лежит в основе поддержания водного баланса у мелких грызунов в некоторых наиболее жарких пустынях земного шара.
| {38} |
В предыдущей главе основной упор делался на то, как трудно животным, особенно мелким, терять воду и тепло из дыхательного тракта. Тем не менее многие млекопитающие и птицы используют испарение из дыхательных путей для выведения из организма излишков тепла. Все мы не раз наблюдали, как в жаркую погоду собака учащенно дышит, высунув наружу язык, — это так называемая тепловая одышка; то же самое делают и другие домашние животные, например овцы и коровы. У птиц при большой тепловой нагрузке также значительно усиливается дыхательная активность, хотя это происходит несколько иначе, чем у млекопитающих, к чему я вернусь ниже.
Тепловую одышку у собак часто описывают как учащенное дыхание через открытую пасть, сопровождающееся испарением с влажных слизистых ротовой полости и с большого свешивающегося наружу языка1. Воздух, протекающий вблизи от слизистых, быстро насыщается водяными парами, однако основная его масса не соприкасается непосредственно с этими влажными поверхностями, и нетрудно понять, что полностью насытить влагой весь объем воздуха не так-то легко. Это вызывает необходимость перемещения больших масс воздуха, а следовательно, требует дополнительных мышечных усилий, что приводит {39} к повышению затрат энергии, а тем самым — и тепловой нагрузки.
A priori создаётся впечатление, что слизистые носа лучше приспособлены для испарения, поскольку воздух теснее соприкасается с обширными влажными поверхностями носовой полости и быстро достигает полного равновесия как в отношении температуры, так и в отношении содержания водяных паров (Верцар, Кейт и Парчет, 1953). Однако отдавать тепло через нос затруднительно, потому что большая часть тепла и водяных паров, добавляемых к выдыхаемому воздуху, вновь возвращаются организму в результате противоточного обмена при выдыхании. Один из способов избежать этого возврата тепла заключается в том, чтобы воздух протекал через нос лишь в одном направлении (снаружи внутрь), а выдыхался через рот.
Оказалось, что у собак при тепловой одышке воздух действительно течет главным образом в одном направлении: вдыхание происходит преимущественно через нос, а выдыхание — через рот (рис. 16). У обследованных нами собак в среднем примерно четвертая часть воздуха, вдыхаемого через нос, выдыхалась также через нос, а остальные три четверти через рот. Это соотношение, однако, сильно колеблется, и в каждый отдельный момент через нос может выдыхаться от 0 до 100% воздуха (Шмидт-Нильсен, Бретц и Тэйлор, 1970, а).
Описанные особенности движения воздуха при тепловой одышке у собак означают, что испарение происходит главным образом со слизистой носа, а не ротовой полости и языка. Соответственно нос должен получать достаточные количества влаги, и мы считаем, что основным ее поставщиком служит крупная носовая железа. Эта железа была описана в 1664 году датским анатомом Николаусом Стено, но поскольку у человека аналогичной железы нет, она не привлекала особого внимания и осталась сравнительно малоизученной. Никаких специальных функций, за исключением «поддержания носа во влажном состоянии», для этой железы не обнаружено; если она действительно поставляет воду, необходимую для теплоотдачи во время тепловой одышки, то ее функция {40} в некотором смысле аналогична функции потовых желез у человека. Поэтому интересно было бы выяснить, не находится ли секреторная функция этой
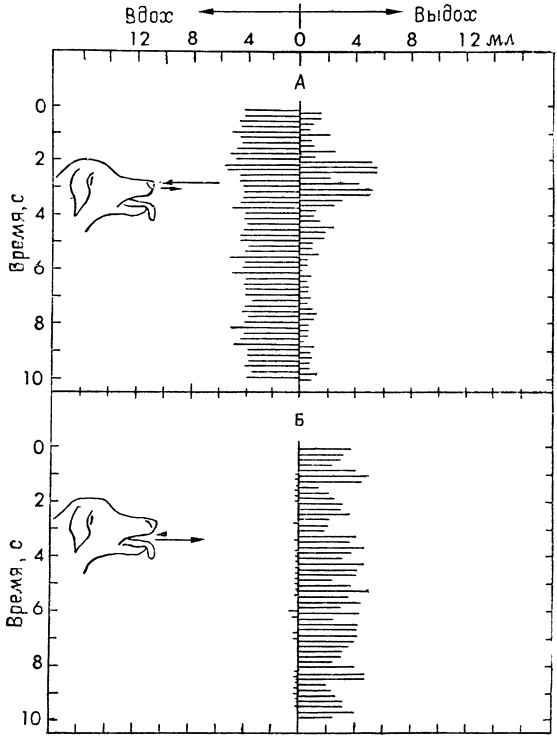 |
|
Рис. 16. Пути прохождения воздуха при тепловой одышке у собаки (по Шмидту-Нильсену и др., 1970, а). А. Поток воздуха через нос. Горизонтальными линиями, отходящими влево от вертикальной средней линии, показан вдыхаемый воздух, а отходящими вправо — выдыхаемый. Средние объемы вдыхаемого и выдыхаемого воздуха обозначены стрелками у носа собаки. Б. Поток воздуха через рот. Обозначения те же. Вдыхание через рот практически равно нулю, но почти весь воздух, вдыхаемый через нос, выдыхается через рот. |
железы, подобно секреции потовых желез, под контролем системы терморегуляции.
О важном значении того, что воздух проходит через нос лишь в одном направлении, можно судить по {41} температуре выдыхаемого воздуха. При умеренной тепловой нагрузке тепловая одышка у собаки происходит при закрытом рте, то есть воздух и вдыхается, и выдыхается через нос; температура выдыхаемого воздуха в этих условиях равна 29°С. Если эта же самая собака, продолжая, тепловую одышку с той же частотой, начнет выдыхать воздух через рот, его температура будет почти равна температуре тела, то есть 38°С. Когда выдыхаемый воздух имеет более высокую температуру, он уносит с собой больше тепла и влаги, усиливая тем самым рассеивание тепла. Один литр насыщенного влагой воздуха, выдыхаемого при температуре 38°С, уносит с собой 27,7 кал тепла, а при 29°С — только 14,9 кал. Таким образом, при выдыхании через рот теплоотдача увеличивается почти вдвое. Изменяя соотношение объемов воздуха, выдыхаемого через нос и через рот, собака может регулировать количество рассеиваемого тепла, не изменяя частоты дыхания или дыхательного объема.
Это подводит нас к интересному факту: оказывается, собаки (и многие другие животные) при тепловой одышке дышат с относительно постоянной частотой. Как только начинается тепловая одышка, частота дыхания внезапно резко повышается, скажем от 30—40 дыхательных циклов в минуту до 300—400. При очень небольшой тепловой нагрузке у собак наблюдается не тепловая одышка с какими-либо промежуточными, частотами, а чередование коротких периодов тепловой одышки при высокой частоте дыхания с периодами нормального дыхания.
В 1962 году Ю. Кроуфорд, в то время студент Университета Дьюка, высказал предположение, что при тепловой одышке собаки дышат с резонансной частотой. Благодаря своим упругим свойствам вся их дыхательная система имеет некоторую собственную частоту колебаний, и для того чтобы поддерживать такое колебание системы, достаточно минимальных мышечных усилий. Поэтому теплопродукция мышц, участвующих в дыхательных движениях, незначительна и очень мало влияет на тепловую нагрузку. Кроуфорд считает, что если бы тепловая одышка происходила в отсутствие такого резонансного режима, то связанное с этим усиление мышечной деятельности {42} привело бы к генерации большего количества тепла, чем может рассеяться в результате тепловой одышки.
Если тепловая одышка происходит при некоторой постоянной резонансной частоте, то изменение количества рассеиваемого тепла не должно сопровождаться изменением этой частоты1. Вот теперь становится понятным, какую важную роль играет способность собак регулировать потери тепла, направляя потоки воздуха то через нос, то через рот.
Хорошо регулируемое изменение потока воздуха через нос дает возможность точно регулировать потери тепла: 1) не изменяя частоты дыхания, что потребовало бы больших затрат энергии и вызвало бы дополнительную теплопродукцию в мышцах; 2) не изменяя глубины дыхания, что сильно сказалось бы на газообмене в легких.
Тепловая одышка при умеренных тепловых нагрузках не приводит к гипервентиляции легких (Элберс, 1961), но при значительных тепловых нагрузках она увеличивает объем легочной вентиляции в минуту, и такая гипервентиляция постепенно ведет к чрезмерным потерям кровью двуокиси углерода и в конечном счете — к алкалозу. Хэйлс и Блай (1969) наблюдали у собак непосредственно после тепловой нагрузки 18-кратное повышение частоты дыхания — от 18 до 340 дыхательных движений в минуту. С дальнейшим увеличением тепловой нагрузки частота дыхания достигла максимума — 410 дыхательных движений в минуту, но затем вновь снизилась до 260. В течение этой второй фазы объем легочной вентиляции в минуту и альвеолярная вентиляция сильно увеличились и возник респираторный алкалоз.
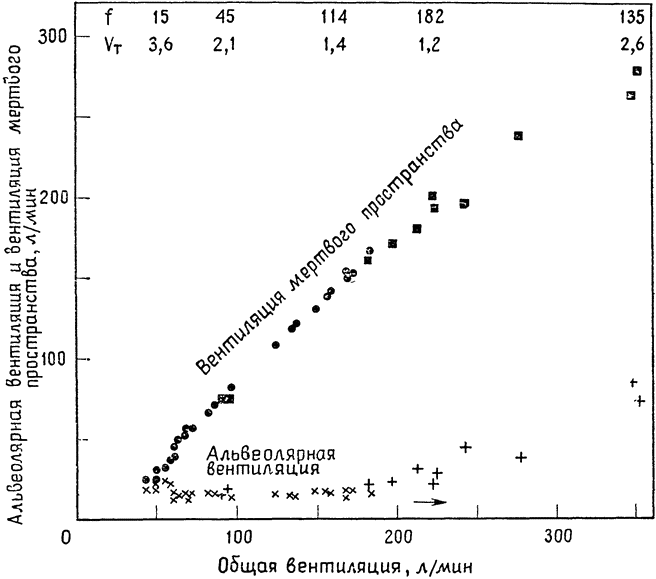
Разделение тепловой одышки на две фазы наблюдается также у овец и быков (Хэйлс и Финдлей, 1968). У быка в течение первой фазы вентиляция мертвого пространства может во много раз увеличиться, не оказывая заметного влияния на альвеолярную вентиляцию (рис. 17); и только при стрессовых тепловых {43} нагрузках, когда дыхание становится затрудненным, животное хватает воздух ртом и проявляет явные признаки беспокойства, альвеолярная вентиляция начинает возрастать. Во время этой второй фазы общая вентиляция легких продолжает возрастать, и несмотря
 |
|
Рис. 17. Легочная вентиляция при тепловой одышке у быка (по Хэйлсу, 1966). По мере увеличения общей вентиляции постепенно возрастает вентиляция мертвого пространства Альвеолярная же вентиляция не возрастает до тех пор, Пока общая вентиляция не превысит 200 л/мин. При крайне выраженной одышке частота дыхания (f) уменьшается с увеличением дыхательного объема (vt) (цифры вверху). |
на то, что это сопровождается снижением частоты дыхания, у животного наблюдается резко выраженный алкалоз.
У большинства птиц при тепловой нагрузке возникает тепловая одышка, выражающаяся в заметном учащении дыхания; у некоторых птиц, кроме того, {44} наблюдается быстрое трепетание горла. При обсуждении особенностей дыхания птиц при высоких температурах среды и способов, которыми они пользуются для охлаждения, не всегда проводится достаточно четкая граница между тепловой одышкой и трепетанием горла. Под тепловой одышкой я здесь имею в виду дыхательные движения груди и брюха, тогда как трепетание горла — это быстрые движения тонкого дна ротовой полости и верхней части шеи. Я почти не буду касаться последнего, а остановлюсь главным образом на тепловой одышке. Явление тепловой одышки у птиц представляет особый интерес, поскольку их дыхательная система в некоторых отношениях резко отличается от дыхательной системы млекопитающих.
Вкратце дыхательная система птиц состоит из парных легких, в которых происходит газообмен между воздухом и кровью, и из нескольких пар больших тонкостенных воздушных мешков, которые действуют как мехи, способствуя просасыванию воздуха через легкие. Эти мешки бедны кровеносными сосудами и не принимают непосредственного участия в газообмене, что наглядно показывает следующий эксперимент. Если в один из воздушных мешков, предварительно перекрыв все его связи с другими частями дыхательной системы,-вводить окись углерода, она не поступаете кровь в количествах, достаточных, чтобы отравить птицу.
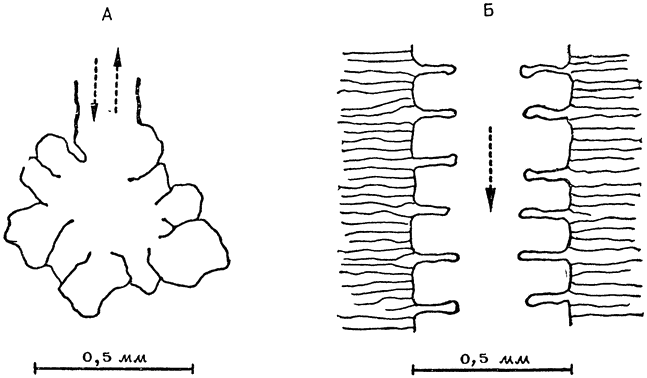
У млекопитающих легкие вентилируются за счет потоков воздуха в двух противоположных направлениях: снаружи — внутрь и изнутри — наружу. Легкие же птиц устроены таким образом, что воздух проходит через них насквозь: самые тонкие веточки бронхов не кончаются у них слепыми мешочками типа альвеол млекопитающих, а представляют собой открытые на обоих концах трубочки; газообмен происходит в воздушных капиллярах, которые окружают эти трубочки, называемые парабронхами, или третичными бронхами (рис. 18).
Анатомия органов дыхания птиц изучена очень детально, однако вопрос о том, как именно протекает воздух по различным воздухоносным путям легкого, остается открытым. На этот счет высказывались многочисленные гипотезы и было опубликовано несколько {45} хороших обзоров, среди которых следует упомянуть наиболее полный и ясный обзор А. С. Кинга (1966).
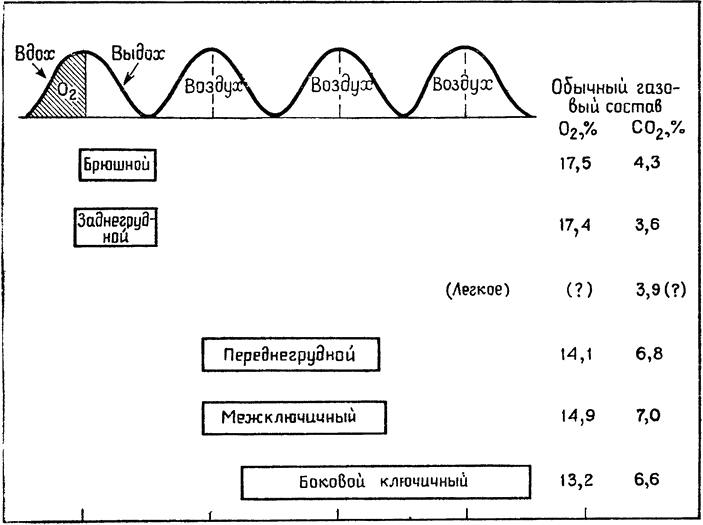
Многие читатели, вероятно, достаточно знакомы со строением дыхательной системы птиц; и тем не менее мне хочется напомнить некоторые наиболее важные для нашего изложения моменты. Вся система воздухоносных путей занимает около 15% объема тела птицы; сами легкие относительно невелики и довольно мало растяжимы, а большую часть системы составляют воздушные мешки (рис. 19), которые проникают своими
 |
|
Рис. 18. Строение самых мелких подразделений легкого у млекопитающих (А) и птиц (Б). У млекопитающих самые мелкие бронхи оканчиваются слепыми мешочками — альвеолами, а у птиц самые тонкие веточки бронхов (парабронхи) представляют собой трубочки, открытые на обоих концах и пропускающие воздух насквозь. |
разветвлениями в кости, создавая так называемую пневматизацию костей, и в другие части тела.
Наиболее своеобразная особенность строения системы органов дыхания птиц — это сложность воздухоносных путей, соединяющих разные части этой системы. Крупный главный бронх, называемый средним бронхом (или мезобронхом), проходит через все легкие, отдавая ветви в передние воздушные мешки; он связан с системой тонких бронхов, пронизывающих легочную паренхиму, и продолжается за пределами легкого, переходя в задние воздушные мешки. Кроме того, воздушные мешки связаны непосредственно {46} с легкими при помощи системы так называемых экто- и энтобронхов, что создает множество дополнительных каналов и направлений, по которым может протекать воздух.
В схематической форме эти основные связи представлены на рис. 20, где все воздушные мешки, которые расположены в передней части тела птицы, объединены в одно пространство, а все мешки, расположенные
 |
|
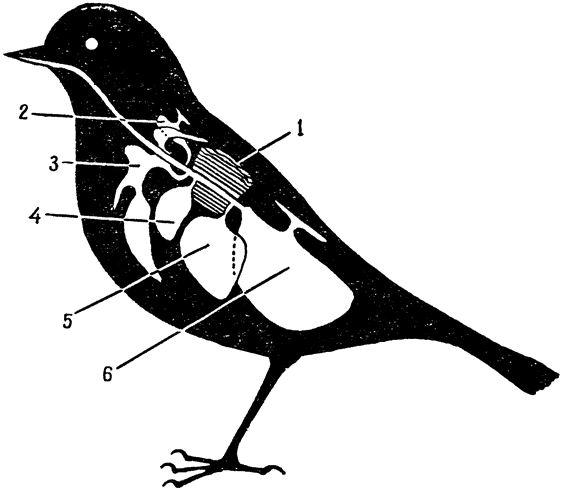
Рис. 19. Схема расположения воздушных мешков в теле птицы (по Солту, 1964). 1 — легкое; 2 — шейный мешок; 3 — межключичный мешок (непарный); 4 — переднегрудной мешок; 5 — заднегрудной мешок; 6 — брюшной мешок |
в задней его части, — в другое; между ними находится легкое. Показаны также воздухоносные пути, связывающие воздушные мешки с легкими. Наиболее важное значение непосредственной связи между крупным средним бронхом и воздушными мешками состоит в том, что она, по-видимому, создает возможность для усиленной вентиляции одних лишь воздушных мешков, не затрагивающей само легкое. Иными словами, имеется некий обходной путь, или шунт, который допускает усиление потока воздуха при тепловой {47} одышке, и притом таким образом, что это не сопровождается увеличением объема воздуха, прогоняемого, через легкое, — оптимальное решение проблемы, позволяющее избежать алкалоза.
Для нашего дальнейшего обсуждения важно указать, что, судя по имеющимся наблюдениям, у многих птиц для тепловой одышки характерна довольно постоянная частота дыхания. Было высказано предположение, что это связано с существованием у них некоторой резонансной частоты, подобной той, которую Кроуфорд обнаружил у собак. Например, когда у голубя начинается тепловая одышка, частота дыхательных движений у него резко изменяется, повышаясь от
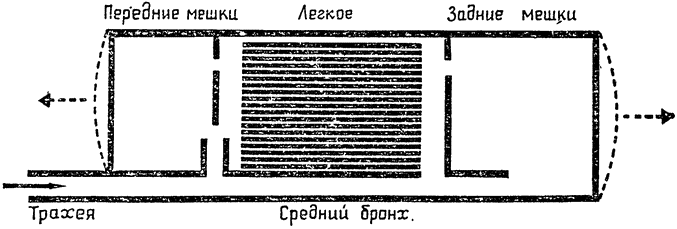 |
Рис. 20. Схема связей между бронхами, легкими и воздушными мешками у птиц. |
30 до примерно 600 циклов в минуту1. Недавно Кроуфорд и Кемпе (1971) показали, что резонансная частота дыхательной системы голубей (564 цикла в минуту) практически равна частоте дыхания при тепловой одышке (612 циклов в минуту). Следовательно, работа, которую нужно затратить на преодоление сил, противодействующих дыхательным движениям, при этой частоте минимальна. Это очень важное открытие, поскольку оно означает, что тепловая одышка обеспечивает птице достаточную для повышения испарения вентиляцию при незначительных затратах энергии (и тем самым лишь минимально увеличивает теплопродукцию мышц, участвующих в дыхании). {48}
Необходимо отметить, однако, это это ни в коей мере не относится ко всем птицам. Например, у бакланов повышение температуры тела сопровождается непрерывным и равномерным повышением частоты дыхания, хотя трепетание горла у этих птиц всегда происходите высокой и постоянной частотой, заставляя предполагать существование здесь специальной колебательной системы (Лазиевский и Снайдер, 1969). У бакланов и пеликанов частоты дыхания и трепетания совершенно независимы; однако у голубей дыхание при одышке и трепетание совершенно синхронны (Кальдер и Шмидт-Нильсен, 1966). Это относится также ко многим другим птицам: гусям, уткам, курам, чайкам, земляным кукушкам и пр. Следует отметить, что у некоторых птиц, в частности у грифов и воробьиных, тепловая одышка не сопровождается ни малейшими признаками трепетания горла.
У птиц, у которых частота дыхания при тепловой одышке определяется резонансными характеристиками дыхательной системы, эта частота остается постоянной, и для регуляции испарения влаги и рассеивания тепла им приходится находить другие способы. Самый простой способ, пригодный при небольших тепловых нагрузках, — это чередование коротких периодов тепловой одышки с периодами нормального дыхания, как это делают собаки. Многие птицы и в самом деле ведут себя таким образом: при небольших тепловых нагрузках у них наблюдается периодическая тепловая одышка. Если же тепловая нагрузка велика, то одышка происходит непрерывно, а дальнейшее усиление испарения и теплоотдачи достигается путем увеличения суммарного потока воздуха, проходящего через дыхательную систему (минутный объем). При этом, вероятно, начинает действовать упоминавшийся выше морфологический шунт, препятствующий прохождению всей массы воздуха через легкие и тем самым предотвращающий алкалоз, который наступает у млекопитающих при сильной тепловой одышке.
Велико же было наше удивление, когда мы обнаружили, что у голубей при сильной тепловой одышке {49} развивается тяжелый алкалоз. Очевидно, шунтирования, если оно и существует, недостаточно для предотвращения гипервентиляции легких (Кальдер и Шмидт
 |
|
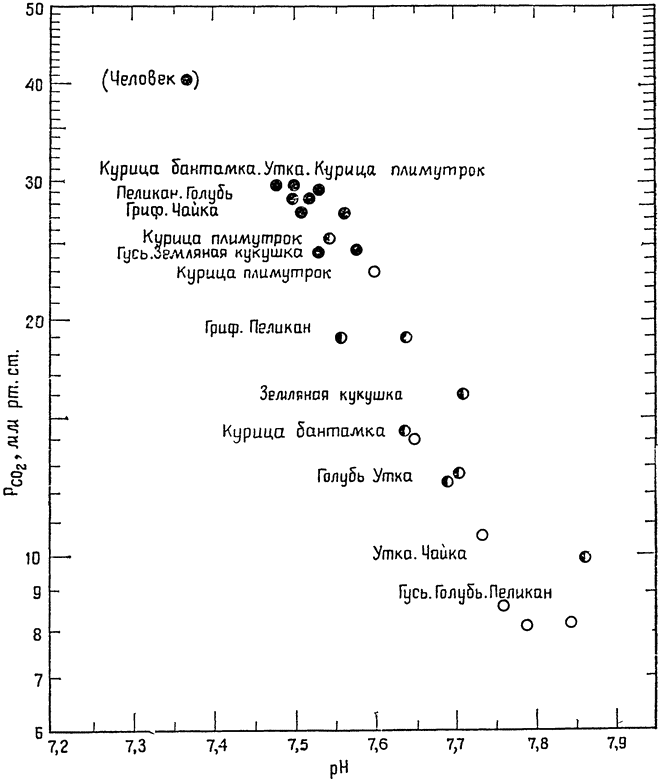
Рис. 21. Напряжение СO2 в артериальной крови птиц на отдыхе (черные кружки) равно примерно 30 мм рт. ст. или чуть ниже (у млекопитающих — 40 мм рт. ст.). При тепловой одышке напряжение СO2 ниже (полузачерненные кружки); при максимальной тепловой одышке (белые кружки) напряжение СO2 может снижаться до 8 мм рт. ст., а рН крови достигает 7,9. Все точки представляют собой средние значения для данного вида (по Кальдеру и Шмидту-Нильсену, 1968). |
Нильсен, 1966). В дальнейшем оказалось, что та же самая картина наблюдается и у некоторых других видов птиц: тепловая одышка сопровождается понижением давления СO2 в крови и таким сдвигом рН крови {50} в щелочную сторону, который для человека оказался бы летальным (рис. 21).
Было бы, однако, ошибкой делать вывод, что у всех птиц при одышке развивается сильный алкалоз. Страус, обитающий в жарких засушливых районах Африки и Ближнего Востока, обладает большой выносливостью к тепловым нагрузкам — в наших экспериментах он выдерживал температуру 56°С в течение 5 часов; на протяжении всего этого времени у него наблюдалась тепловая одышка с частотой дыхания 50 циклов в минуту. Объем воздуха, проходящего через дыхательную систему страуса, может при тепловой одышке возрасти в 20 раз по сравнению с тем Объемом, который проходит через эту систему, когда птица отдыхает. Газовый состав воздуха, находящегося в воздушных мешках, при тепловой одышке приближается к составу наружного воздуха и содержание в нем СO2 снижается до менее чем 1%. Это свидетельствует об интенсивной вентиляции воздушных мешков, но тем не менее алкалоза не возникает (Шмидт-Нильсен и др., 1969). Таким образом, мы вправе предположить, что у страуса морфологический шунт действует в полную силу. Остается загадкой, почему у других птиц все происходит иначе. Вряд ли можно связать это различие с тем, что страусы не летают, поскольку у них имеется вполне развитая система воздушных мешков, сходная с таковой у других птиц.
| {51} |
Неожиданное открытие, что большинство птиц при тепловой одышке, по-видимому, не используют явно имеющийся у них морфологический шунт, чтобы направлять поток воздуха в обход легкого, делает еще более желательным выяснение путей, по которым протекает воздух в их сложной дыхательной системе.
В дыхательной системе млекопитающих нет ничего сложного: мешковидные легкие и их доли вентилируются двумя встречными потоками воздуха, а газообмен в концевых элементах — альвеолах — происходит главным образом при помощи диффузии. Птицам же не нужен поток воздуха в обратном направлении, так как самые тонкие веточки бронхов — парабронхи, или третичные бронхи, — открыты на обоих концах, так что воздух проходит через них насквозь. Эта особенность в сочетании с наличием у птиц множества трубок, веточек и соединений с воздушными мешками породила ряд гипотез о том, как именно протекает воздух по столь сложной системе. Высказывались всевозможные предположения, в том числе и такие, которые противоречат физическим законам, наблюдаемым фактам и данным физиологии. Поэтому мы начнем с краткого обзора имеющихся в настоящее время данных, довольно убедительно указывающих на то, что через легкие птиц воздух течет в одном направлении как при вдохе, так и при выдохе, а затем обсудим, какое это имеет значение для функции легких в качестве органа поглощения кислорода. Хотя кровь птицы обладает не большим сродством к кислороду, чем кровь млекопитающего, строение птичьего легкого обеспечивает гораздо более полное, чем у млекопитающих, использование кислорода при низком атмосферном давлении, что очень важно при полете на больших высотах. {52}
Необходимость более глубокого понимания механизма дыхания птиц приобрела особую остроту в процессе изучения этой функции у страуса: как показывали полученные результаты, воздух протекает по его легким в одном (краниальном) направлении и при вдохе, и при выдохе.
Интересно отметить, что у страуса, хотя он и не летает, имеется хорошо развитая система воздушных мешков, сходная по общему строению с таковой у других птиц и составляющая примерно 15% объема его тела (Шмидт-Нильсен др., 1969). Для того чтобы выяснить, как протекает воздух по этой системе, страусу давали вдохнуть один раз чистый кислород (в качестве метки), а затем с помощью кислородного электрода регистрировали время его поступления в разные пункты системы воздушных мешков (рис. 22). Обычно некоторое количество вдыхаемого кислорода доходило до задних мешков к концу первого вдоха или сейчас же после него. Содержание кислорода в задних мешках продолжало возрастать в течение всей фазы выдоха, однако в течение второго цикла дыхания дальнейшего возрастания не наблюдалось. Тем самым подтверждается предположение, что вдыхаемый газ (во втором цикле это был снова атмосферный воздух) поступает прямо в задние воздушные мешки.
Поступление кислорода в передние мешки всегда происходило с некоторой задержкой — не раньше, чем во время второго или третьего цикла дыхания. Очевидно, тот кислород, который в конечном счете появлялся в передних мешках, сначала попадал куда-то еще. Мы пришли к выводу, что воздух, поступающий в передние мешки, приходит в них из задних мешков и (или) из легких, и сформулировали свое заключение следующим образом: «Вдыхаемый воздух поступает сначала в задние мешки (хотя некоторая его часть может непосредственно проходить в легкие); затем, во время выдоха, большая часть воздуха из задних мешков поступает в легкие, и таким образом в передние мешки, приходит воздух, который уже прошел через легкие» (Шмидт-Нильсен и др., 1969).
Низкая частота дыхания у страуса (примерно 6 циклов в минуту) позволяет высказать еще одно {53} интересное предположение: по-видимому, воздушные мешки очень хорошо вентилируются. Используя в качестве метки чистый кислород, мы установили, что время его вымывания как для задних, так и для передних воздушных мешков очень невелико; иными словами, ни в одном из мешков не застаивалась
 |
|
Рис. 22. Время поступления метки (чистый кислород) в разные части системы воздушных мешков у страуса (по Шмидту-Нильсену и др., 1969). Вверху вычерчен дыхательный цикл. Заштрихованная часть первого цикла соответствует вдыханию чистого кислорода. Частота дыхания — примерно 6 циклов в минуту, то есть продолжительность 1 цикла равна примерно 10 с. Левая граница каждого горизонтального прямоугольника соответствует времени первого зарегистрированного появления кислорода в данном мешке, а правая граница — времени последнего зарегистрированного повышения содержания в нем кислорода. Кислород сначала поступает в задние воздушные мешки, а затем, спустя 1—2 цикла, — в передние. |
инертная масса воздуха. Короткое время вымывания для передних мешков, для которых характерно высокое содержание СO2 и относительно низкое содержание O2 (за исключением того времени, когда у птицы происходит тепловая одышка), должно означать, что в эти мешки не попадает наружный воздух. Газовый состав наполняющего их воздуха легче всего объяснить, {54} исходя из предположения, что они наполняются воздухом, поступающим из легких, где он уже находился в контакте с поверхностями, на которых происходит газообмен.
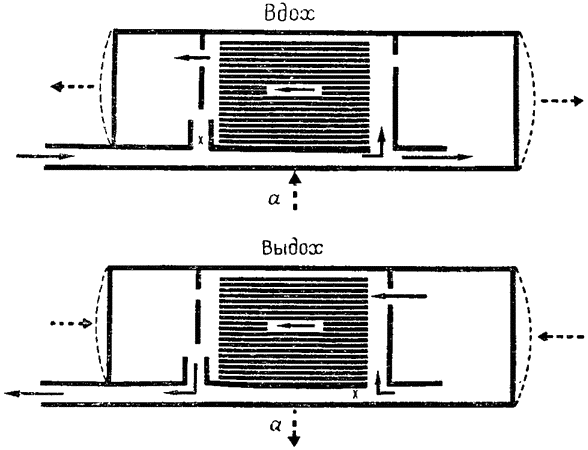
Один из моих сотрудников, В. Бретц, разработал анемометрический датчик, позволяющий регистрировать направление потока воздуха и имеющий достаточно малые размеры, чтобы его можно было ввести в наиболее важные пункты дыхательной системы уток. Для дальнейшего изложения следует вновь вернуться к упрощенной схеме дыхательной системы птиц, изображенной на рис. 20. Напомню, что все передние воздушные мешки представлены на ней в виде единого переднего пространства, а все задние — в виде единого заднего пространства. Главный бронх непосредственно связан с брюшным воздушным мешком, отдавая по пути ветви в передние мешки, а также в сами легкие. Кроме того, воздушные мешки непосредственно соединены с легкими системой экто- и энтобронхов, что увеличивает число отверстий и связей.
Изучение картины потока воздуха в легких птицы позволило построить схему, изображенную на рис. 23 (Бретц и Шмидт-Нильсен, 1970, 1971). При вдохе как передние, так и задние мешки растягиваются1. Поток наружного воздуха направляется главным образом в задние мешки, которые таким образом получают смесь воздуха из мертвого пространства легких и свежего атмосферного воздуха. Часть вдыхаемого воздуха поступает также в легкие через самое заднее (каудо-дорсальное) отверстие. У утки, как и у страуса, при вдохе атмосферный воздух, очевидно, в передние воздушные мешки не поступает; следовательно, они должны наполняться воздухом из легких. Заключение о том, что наружный воздух не поступает {55} в передние мешки, подтверждается результатами анализа газового состава воздуха в этих мешках, а также тем, что анемометрический датчик, помещенный в месте их соединения с главным бронхом (указано буквой х на рис. 23), не зарегистрировал потока воздуха. Наличие потока воздуха, направленного из легких в передние мешки, — логическое следствие, согласующееся с нашими наблюдениями на страусах. Поскольку
 |
|
Рис. 23. Пути воздушного потока в дыхательной системе птиц. Воздух непрерывно проходит через легкие как при вдохе, так и при выдохе. Стрелками а показано направление движений легочного апоневроза. |
мешки действительно растягиваются, то единственной причиной этого может быть воздух, поступающий из легких. Справедливость подобного заключения подтверждается высоким (около 6%) содержанием СO2 в передних мешках.
Во время выдоха (рис. 23) поток воздуха в среднем бронхе (помечен буквой х) незначителен, так что задние воздушные мешки должны опорожняться в само легкое. Передние же мешки опорожняются непосредственно в начальную часть бронха, как показывает довольно интенсивный поток воздуха в соответствующих бронхиальных соединениях. {56}
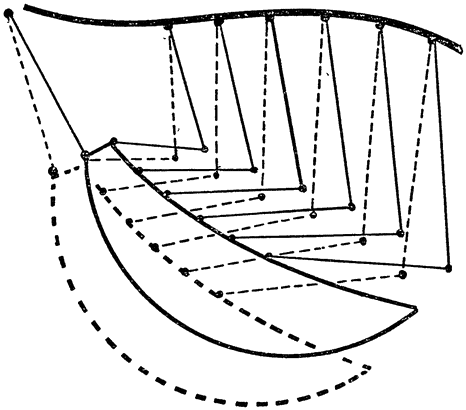
Предположение о том, что при выдохе воздух течет из задних мешков в легкие, наталкивается на одно затруднение. Принято считать, что легкое птицы относительно мало растяжимо и что его объем на протяжении дыхательного цикла изменяется очень незначительно. С подобным представлением трудно согласиться. Строение грудной клетки птиц с ее подвижными сочленениями между спинными и брюшными частями ребер как бы специально приспособлено для
 |
|
Рис. 24. Строение скелета грудной клетки птиц. Подвижное сочленение спинного и брюшного отделов каждого ребра позволяет грудине при дыхании то приближаться к позвоночнику, то удаляться от него. При этом объем грудной клетки, в которой находятся легкие и некоторые из воздушных мешков, сильно изменяется. |
больших изменений ее объема (рис 24). Обычно считается, что прохождение воздуха в легких у птиц происходит за счет того, что воздушные мешки выполняют роль мехов. Это, казалось бы, подтверждается отсутствием у птиц мышечной диафрагмы; однако у них имеется апоневроз — мембранная структура, располагающаяся вдоль брюшной поверхности легких. Апоневроз слегка выгнут и несколько вдается в легкие, так что при его сокращении они должны растягиваться. Бретц недавно высказал предположение, что мышцы, связанные с апоневрозом, участвуют в движении воздуха из задних мешков в легкое при выдохе.
| {57} |
Рис. 25. Продольный разрез легкого утки. Обратите внимание на толстые вторичные бронхи, отходящие от задней части среднего бронха. Эти вторичные бронхи, заворачивающиеся на спинную поверхность легкого, имеют очень тонкие стенки и, несомненно, должны растягиваться при давлении на брюшную поверхность легких со стороны апоневроза (толстая линия внизу).
| {58} |
Хотя растяжимость самого легкого невелика1, имеющиеся на его спинной поверхности многочисленные крупные вторичные бронхи могут легко растягиваться под действием мышц, оказывающих давление на апоневроз (рис. 25). В связи с этим интересно вспомнить, что Соум (1896) наблюдал сокращение апоневроза при выдохе; кроме того, при выдохе была зарегистрирована электрическая активность связанных с апоневрозом мышц (Федде, Барджер и Китчелл, 1963). Таким образом, мы приходим к, казалось бы, парадоксальному выводу: легкие птицы наполняются воздухом при выдохе и опорожняются при вдохе.
Многие авторы подчеркивали, что в дыхательной системе птиц нет каких-либо клапанов, которые бы определяли направление потока воздуха. Чисто теоретически меня больше всего смущало то обстоятельство, что при падении давления в передних мешках во время вдоха воздух из главного бронха в эти мешки, по-видимому, не поступает. Этот вывод, очевидно, можно считать правильным, поскольку специальные датчики не регистрировали здесь никакого воздушного потока; кроме того, при вдыхании индикаторных газов они не поступали непосредственно в передние мешки.
В целом дыхательная система птиц — это высокоскоростная система низкого давления, поэтому для направленного потока воздуха важную роль играют аэродинамические характеристики системы. Отверстия вторичных бронхов, по-видимому, удивительно удачно устроены с гидродинамической точки зрения; вполне возможно, что их форма и ориентация играют важную роль в том, чтобы направлять поток воздуха при вдохе только в главный бронх.
Следует отметить, что сопротивление потоку воздуха со стороны легкого, по-видимому, подвержено резким изменениям. Внутренние стенки парабронхов {59} покрыты тонким сплетением гладких мышечных волокон, которые, сокращаясь, изменяют их диаметр. Сокращение гладких мышц можно вызвать механическим раздражением; их можно также заставить сокращаться или расслабляться под воздействием различных лекарственных веществ (Кинг и Коуи, 1969).
Пытаясь выяснить роль парабронхов в изменении потока воздуха, мы измеряли изменение их диаметра
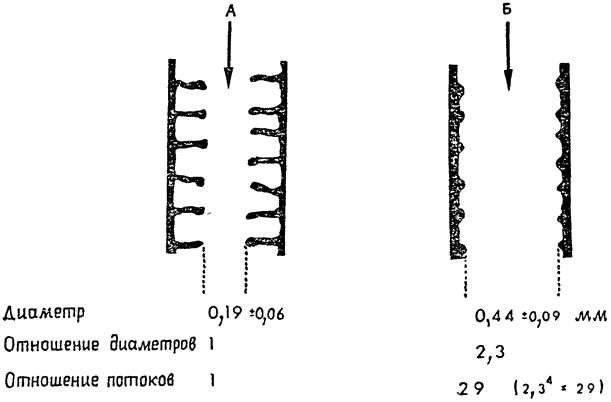 |
Рис. 26. Внутренний диаметр парабронхов из легкого утки уменьшается при воздействии метахолином (A) и увеличивается при воздействии адреналином (Б); сопротивление потоку воздуха изменяется при этом почти в 30 раз. |
в легком утки. Мышцы расслабляли, вводя в них адреналин, а затем заставляли сокращаться, вводя метахолин (холинэргический препарат, удерживающийся в крови позвоночных дольше, чем ацетилхолин). Измерения показали, что диаметр парабронхов в легком утки может изменяться в 2,3 раза (рис. 26). Поскольку объем, протекающий через трубку, пропорционален четвертой степени радиуса (закон Пуазейля), это приводит при постоянном давлении к почти 30-кратному изменению объема. В какой мере подобные изменения способствуют регуляции потока воздуха в дыхательной системе живой птицы — вопрос, изучением которого следует заняться.
| {60} |
Предположение о том, что поток воздуха проходит через легкое птицы только в одном направлении, высказывалось неоднократно. Более того, по мнению некоторых авторов, воздушные мешки служат для обеспечения непрерывного поступления в легкие свежего воздуха извне, с тем чтобы кровь получала максимально возможное количество кислорода. Однако подобное положение вещей было бы совершенно нецелесообразным: оно было бы равносильно чрезвычайно интенсивной гипервентиляции легких и приводило бы к вымыванию из крови больших количеств СO2, а также к резкому нарушению кислотно-щелочного равновесия.
У птиц и млекопитающих для нормального дыхания кровь должна достигать равновесия при довольно высоком содержании СO2. Именно это обеспечивает предложенная нами модель. Задние мешки служат смесительными камерами, в которых происходит перемешивание поступающего при вдыхании атмосферного воздуха с воздухом мертвого пространства; получающаяся в результате смесь содержит 3—4% СO2 (такое содержание СO2 часто обнаруживается при анализе воздуха, наполняющего задние мешки). Эта смесь, все еще богатая O2, проходит через легкие, обеспечивая полное насыщение крови кислородом. Выходящий из легких воздух близок к равновесию с венозной кровью и отличается высоким содержанием СO2 (около 6% и даже больше). Этот воздух поступает в передние мешки, играющие роль сборных камер, из которых воздух выводится наружу при следующем выдохе.
Однонаправленный поток воздуха в легком птицы заставляет подумать о возможности существования здесь противоточной системы между воздухом и кровью. Подобная противоточная система имеется в жабрах рыб, благодаря чему артериальная кровь получает возможность насыщаться кислородом при {61} таком его напряжении, какое имеется в притекающей, а не в вытекающей воде. В легких млекопитающих это невозможно, и напряжение кислорода в артериальной крови в лучшем случае соответствует его напряжению в выдыхаемом альвеолярном воздухе.
В жаберных лепестках рыб вода и кровь текут в противоположных направлениях (рис. 27). В результате кровь, как раз перед тем как она покидает
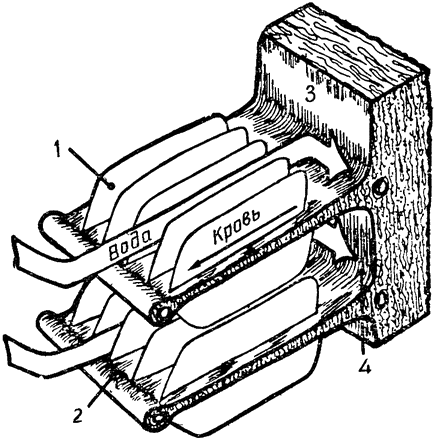 |
|
Рис. 27. В жабрах акулы вода и кровь текут в противоположных направлениях, что создает возможность для большего насыщения крови кислородом (по Григгу, 1970). 1 — вторичные лепесточки; 2 — край жаберного лепестка; 3 — перегородка; 4 — канал. |
жабры, встречается с поступающей снаружи водой, которая еще содержит весь свой запас кислорода; иными словами, напряжение кислорода в крови приблизится к напряжению кислорода в воде, прежде чем вода отдаст какую-то часть кислорода. На другой стороне жаберных лепестков вода, выходящая из жаберной полости, встречается с венозной кровью и, несмотря на то, что вода уже отдала большую часть своего кислорода, кровь извлекает из нее еще некоторое его количество. В результате этого рыбы извлекают из воды до 80—90% содержащегося в ней кислорода; {62} без противоточной системы достигнуть такой эффективности было бы невозможно.
Данные в пользу существования противоточной системы в легких птиц были получены при изучении страуса. У этой птицы РCO2 в артериальной крови ниже, чем в передних воздушных мешках. Много лет назад Зейтен (1942), с которым мы тогда работали в. одной лаборатории в институте профессора Крога в Копенгагене, отметил, что воздух, выходящий из легких, при выходе должен быть гораздо богаче СO2, чем можно было бы думать на основании напряжения СO2 в артериальной крови. Для того чтобы объяснить это, он предложил модель, по которой в конечном счете происходит смешивание порций крови, производящих газообмен с воздухом различного газового состава. Сравнительно недавно Шейд и Пиипер (1970) детально проанализировали эту систему, названную ими «системой перекрестного обмена». Идея о противоточной системе не возникла у Зейтена, вероятно, потому, что он допускал изменение направления потока воздуха на обратное между вдохом и выдохом.
У некоторых видов птиц напряжение СO2 (РCO2) артериальной крови отличается постоянством и находится на сравнительно низком уровне (РCO2 близко к 30 мм или ниже), тогда как в передних воздушных мешках у очень многих птиц оно заметно выше (РCO2 часто выше 40). Такие высокие концентрации CO2 можно было бы ожидать, если бы речь шла о воздухе, застоявшемся в каком-либо замкнутом пространстве; а поскольку воздушные мешки хорошо вентилируются, остается предположить, что высокие значения СO2 — результат газообмена, присходящего каким-то иным способом, отличным от газообмена у млекопитающих. Этот вывод можно распространить на всех птиц, основываясь на работе Лазиевского и Кальдера (1971), которые собрали воедино ценные количественные данные о дыхательных процессах у многих видов птиц. Их хорошо продуманные и документированные выводы сводятся к тому, что «птицы извлекают относительно больше кислорода из вдыхаемого воздуха, чем млекопитающие», а также что напряжение двуокиси углерода в артериальной крови птиц «соответствует примерно 70% такового у млекопитающих». Эти {63} обобщения безусловно верны, однако лежащие в их основе процессы были бы невозможны при газообмене путем диффузии, характерном для млекопитающих; и нам, по-видимому, не остается ничего другого, как предположить наличие у птиц противоточной системы. Однако самым убедительным доводом в пользу такого заключения служит способность птиц летать на высоте более 6000 м.
Дает ли птицам их сложная дыхательная система какие-то преимущества по сравнению с млекопитающими? Мы знаем, что в состоянии покоя как у птиц, так и у млекопитающих наблюдается одинаково высокая интенсивность метаболизма (Лазиевский и Доусон, 1967). Во время полета интенсивность метаболизма у птиц возрастает раз в десять или больше, однако аналогичное повышение потребления кислорода происходит и у человека при тяжелой физической нагрузке. Поэтому мы не можем согласиться с утверждениями о том, что птицам нужно больше кислорода, чем млекопитающим, и что, следовательно, их дыхательная система должна быть более «эффективна». Возможно, птицы действительно более способны выдерживать большие физические нагрузки в течение длительного времени, как, например, при миграционных полетах, однако само по себе это не должно требовать принципиально иной организации дыхательной системы. Об этом свидетельствует также то обстоятельство, что летучие мыши, легкие которых ничем не отличаются от легких других млекопитающих, — превосходные летуны, а некоторые виды совершают даже миграционные перелеты на большие расстояния. Первые определения интенсивности метаболизма у летучих мышей при длительном полете показали, что они потребляют не меньше кислорода (27,5 мл в час на 1 г веса тела), чем летящие птицы (Томас и Сазерс, 1970).
Однако при полете на больших высотах картина резко меняется. Тьюкер (1968, а) из Университета Дьюка, изучая полет птиц, поместил воробьев в условия, {64} имитирующие высоту 6100 м: атмосферное давление 349 мм рт. ст. и парциальное давление кислорода 73 мм. В этих условиях никаких изменений в поведении воробьев не наблюдалось; они были так же проворны и активны, как и в условиях, соответствующих уровню моря, и хотя не начинали летать по собственной воле, но если их осторожно выпускали в воздух, были способны летать и набирать высоту. Между тем мыши, которые на уровне моря находились в состоянии непрерывной активности и обследовали все вокруг, на высоте 6100 м впадали в коматозное состояние, лежали на брюшке и еле ползали.
Сразу же напрашивается вопрос: а не обладает ли кровь воробьев более высоким по сравнению с кровью мышей сродством к кислороду, а тем самым и способностью извлекать из воздуха большие его количества? В начале первой главы мы приводили пример с гуанако, у которого кривая диссоциации кислорода сдвинута влево; это свидетельствует о более высоком сродстве его крови к кислороду, что обеспечивает более высокое насыщение крови кислородом при низком его напряжении на больших высотах в Андах, где обитает это животное. Однако от такого объяснения пришлось отказаться, так как Тьюкер установил, что кривая диссоциации кислорода для крови воробья почти идентична кривой диссоциации для мыши. Исходя из допущения, что газообмен между кровью и воздухом в легких происходит у воробья таким же образом, как и у млекопитающих, Тьюкер вычислил, что артериальная кровь воробьев на высоте 6100 м должна быть насыщена кислородом на 24%. Если же пересмотреть данные Тьюкера на основе допущения, что в легком птицы потоки воздуха и крови направлены в противоположные стороны, то газообмен должен происходить иначе и насыщение артериальной крови воробья кислородом достигает 80% (рис. 28). Эта величина гораздо ближе к тому уровню насыщения кислородом, который необходим млекопитающему, чтобы оставаться в сознании. Поскольку артериальная кровь могла содержать втрое больше кислорода, чем получалось по вычислениям Тьюкера, оценки производимой сердцем работы и ударного объема крови должны быть соответственно снижены. {65}
Итак, в заключение мы можем сказать, что на уровне моря способность дыхательной системы к извлечению кислорода из воздуха у птиц и у млекопитающих одинакова. Однако на больших высотах между ними выявляются резкие различия. Птицам при полете необходимо повышать интенсивность метаболизма примерно в 10 раз, и все же они могут летать
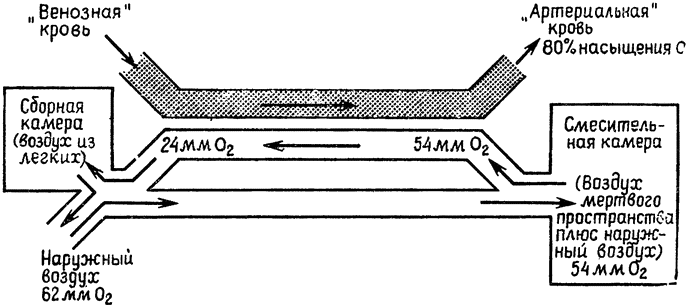 |
|
Рис. 28. Противоположно направленные потоки воздуха и крови (противоточная система) в легких птицы дают возможность артериальной крови прийти в равновесие с богатым кислородом воздухом из задних воздушных мешков. Цифры на схеме соответствуют высоте 6100 м с поправкой на давление водяных паров 54 мм рт. ст. (39,4°С). |
на таких высотах, на которых млекопитающие совершенно неспособны сохранять активность и едва выживают.
Смешивание воздуха мертвого пространства со свежим наружным воздухом, происходящее в задних мешках, имеет одно интересное следствие — величина мертвого пространства с физиологической точки зрения перестает играть какую-либо роль. При этом невероятно длинная трахея, обнаруженная у таких прекрасных летунов, как лебеди и журавли, не столь нелепа, как кажется на первый взгляд. У лебедя-трубача трахея такая длинная, что образует S-образную петлю в грудине (рис, 29). Ее общий объем в 3,3 раза {66} больше, чем следовало бы ожидать для птицы таких размеров (Хайндз и Кальдер, 1971). Подобное увеличение мертвого пространства без видимых на то причин представляется совершенно бессмысленным и побуждает искать какое-либо рациональное объяснение. Попробуем проанализировать эту особенность морфологии лебедя. Обширное мертвое пространство требует соответствующего увеличения объема вдыхаемого
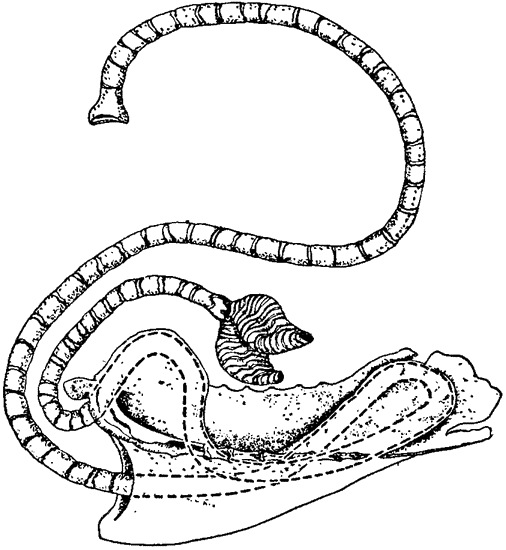 |
Рис. 29. Трахея лебедя-трубача (по Бэнко, 1960). |
воздуха, с тем чтобы при вдохе возникло необходимое соотношение воздуха мертвого пространства и свежего воздуха. При постоянном уровне вентиляции увеличение объема вдыхаемого воздуха означает уменьшение частоты дыхания. И в самом деле, у птиц, как правило, частота дыхания гораздо ниже, чем у млекопитающих, хотя поглощение кислорода у них примерно одинаково: на 1 кг веса тела частота дыхания у птиц более чем вдвое ниже по сравнению с млекопитающими (Лазиевский и Кальдер, 1971). У крупных птиц взмахи крыла и дыхание, возможно, {67} синхронизированы (Тьюкер, 1968, б). Это предположение высказывалось довольно давно просто на основании изучения анатомии птиц; мы не располагаем соответствующими данными для достаточно большого числа видов и поэтому вполне возможно, что нижеследующие рассуждения придется в будущем пересмотреть. Во всяком случае, по чисто аэродинамическим соображениям частота взмахов крыла для данного вида птиц остается довольно постоянной, даже при изменении скорости полета. Если дыхание во время полета синхронизировано со взмахами крыльев, то надо полагать, что именно частота взмахов определяет частоту дыхания; объем вдыхаемого воздуха должен соответствовать низкой частоте дыхания, и в конечном счете для достижения подходящей концентрации СO2 в задних воздушных мешках (в смесительной камере) оказывается необходимым обширное мертвое пространство. Мы знаем, что такие крупные птицы, как лебеди и журавли, машут крыльями очень медленно, и именно у этих птиц обнаружено, казалось бы, бессмысленное увеличение мертвого пространства, выражающееся в необычайно длинной трахее. Подобная интерпретация может оказаться ошибочной, однако она представляется не более рискованной, чем утверждение, что длинная трахея способствует вокализации: ведь многие птицы прекрасно поют, хотя длина трахеи у них гораздо ближе к «нормальной».
| {68} |
В этой главе мы рассмотрим затраты энергии при различных видах активности животного, влияние активности на тепловой баланс и необходимость испарения влаги для предотвращения перегрева.
В конструируемых человеком самолетах двигатели для экономии веса охлаждаются воздухом. Ни одному авиаконструктору не придет в голову нагружать самолет водой, чтобы при длительных перелетах охлаждать мотор за счет испарения. Тем не менее было принято считать, что, поскольку птицы вырабатывают при полете очень много тепла, им для охлаждения своего тела необходимо использовать воду. Было вычислено, что голубю, летящему со скоростью 70 км/час, требуется для охлаждения по 8 г воды на каждый грамм жира, расходуемого в качестве топлива. Вероятно, эти расчеты основаны на неверных допущениях относительно продукции и рассеивания тепла птицей во время полета. Это становится ясно, если мы рассмотрим миграции птиц, при которых они совершают перелеты на огромные расстояния — на многие сотни, а может быть, и тысячи километров, причем иногда они летят над открытым морем, где нет никакой возможности получить воду или корм (Современный, хорошо продуманный обзор по энергетике миграций птиц дает Тьюкер, 1917). К началу перелета до 50% веса птицы составляет жир. Было бы совершенно нелепым полагать, что она, кроме того, несет в себе воду, вес которой должен был бы в несколько раз превышать вес этого жира. {69}
Имеется довольно много сообщений о том, что организм мигрирующих птиц к концу длительного перелёта оказывается сильно обезвоженным, однако нам кажется, что авторы этих сообщений принимали желаемое за действительное; во всяком случае, они не приводят убедительных доказательств в пользу своих утверждений. Напротив, результаты исследования трупов птиц, которые погибают при миграциях, ударяясь о телевизионные, башни и другие препятствия, доказали, что никакого серьезного обезвоживания не происходит, хотя во многих, случаях такие птицы уже пролетели большие расстояния и исчерпали свои запасы жира (см., например, Одум, Роджерс и Хикс, 1964). В одном из присланных мне писем Одум подвел итоги своим многолетним исследованиям в следующих словах: «Энергия, а не обезвоживание является фактором, ограничивающим продолжительность миграционных полетов. Содержание воды, отнесенное к общему весу тела, за вычетом жира, остается у мигрирующих птиц почти постоянным независимо от этапа миграции».
Если бы мы захотели сконструировать птицу с воздушной системой охлаждения, с чего бы мы начали? Эту проблему удобнее обсудить в общем плане, а не применительно к полету. Прежде всего мы придали бы птице высокую температуру тела, что облегчило бы отдачу тепла в окружающую среду без участия испарения. В норме температура птиц равна 40—42°С, то есть она достаточно высока, чтобы допускать теплоотдачу без испарения, если только речь не идет о каких-либо крайних климатических условиях.
Таким образом, при желании мы могли бы рассматривать относительно высокую температуру тела птиц как фактор, способствующий уменьшению использования воды для охлаждения, или, иными словами,— уменьшению необходимости нести на себе лишний груз при полете. Стремление к экономии воды связано не только с полетом. Мелкие грызуны, обитающие в областях с жарким климатом, избегают необходимости расходовать воду на охлаждение тела, прячась в подземные норы. Среди птиц лишь очень немногие живут в норах, так как высокая температуpa {70} тела позволяет им экономить воду, которую, пришлось бы затрачивать на охлаждение. (Многие крупные птицы, например грифы, спасаются от полуденного зноя, паря высоко в небе, где температура ниже.) Поскольку потери воды, порождаемые испарением с поверхности дыхательных путей, неизбежны, приходится прибегать также к другому способу ее экономии — к повышению количества кислорода, извлекаемого из вдыхаемого воздуха. Это позволяет понизить суммарный вентиляционный объем, необходимый при данной скорости потребления кислорода. Как уже говорилось в предыдущей главе, однонаправленный поток воздуха в легких птицы создает возможность для извлечения больших количеств кислорода из выдыхаемого воздуха без повышения сродства крови к кислороду. Вполне возможно, что у птиц во время полета количество извлекаемого из воздуха кислорода повышается настолько, что количество воды, испаряющейся из дыхательных путей, оказывается меньше количества образующейся за то же время окислительной воды: иными словами, может статься, что миграционный полет не требует дополнительных затрат запасов воды. На больших высотах дело обстоит, возможно, несколько иначе, так как при низком парциальном давлении кислорода для получения тех же его количеств вентиляционный объем должен быть выше. Вместе с тем низкая температура воздуха на больших высотах способствует уменьшению испарения при дыхании, хотя мы и не знаем, в какой степени это может компенсировать ожидаемое увеличение.
На протяжении многих лет терморегуляцию и ее влияние на испарение влаги изучали на животных, которых подвергали действию высоких температур в разного рода камерах и помещениях, подогреваемых извне. При этом главное внимание уделялось тепловой нагрузке, создаваемой этим внешним теплом, а повышение «внутренней» тепловой нагрузки, которое вызывается активностью самого организма, изучалось относительно редко, если не считать исследований на человеке и нескольких работ на собаках. Поэтому {71} мы занялись исследованием активности животных и ее влияния на терморегуляцию.
Начали мы с того, что определили для ряда млекопитающих (от мыши до собаки средних размеров) повышение потребления кислорода, а тем самым и теплопродукции при беге. Оказалось, что потребление кислорода возрастает в линейной зависимости от скорости бега, причем наклон кривой для разных видов
 |
Рис. 30. Изменение интенсивности метаболизма в зависимости от скорости бега у животных разных видов (по Тэйлору, Шмидту-Нильсену и Раабу, 1970). |
различен (рис. 30). Прирост потребления кислорода при небольшом ускорении бега выражается наклоном линий регрессии. Этот наклон отражает зависимость потребления кислорода (в мл O2 в час на 1 г веса тела) от скорости бега (в км/час). Если продолжить эти прямые до пересечения с осью ординат, что соответствует нулевой скорости бега, то мы получим на оси ординат некоторый отрезок; вычитая его величину из величины потребления кислорода при любой данной скорости бега, получим значение чистой затраты {72} кислорода на бег при этой скорости. Вычисленное таким образом количество кислорода, которое необходимо затратить на пробег определенного расстояния, оказывается постоянным независимо от скорости бега1.
Среди изученных нами видов затраты кислорода на движение оказались гораздо выше у мелких, чем у крупных животных (рис. 31). То обстоятельство, что-крупному животному перенос 1 г веса на расстояние 1 км с энергетической точки зрения обходится дешевле, чем мелкому, вероятно, можно рассматривать как одно из эволюционных преимуществ больших размеров тела; между тем этому преимуществу, насколько мне известно, как правило, не придают значения.
Мы изучали животных разных размеров — от мыши до собаки. После того как мы вычислили линию регрессии, наилучшим образом соответствующую полученным точкам, оказалось, что имеющиеся в литературе аналогичные данные для овцы и лошади удивительно хорошо ложатся на продолжение этой линии. Интересно отметить, что данные для человека неизменно располагались примерно вдвое выше, чем следовало бы ожидать для четвероногого животного соответствующего веса. Почему же человек на перенос {73} 1 г своего веса на расстояние 1 км расходует вдвое больше энергии, чем можно было ожидать? Объясняется ли это отклонение различием между передвижением на двух или четырех ногах? А если так, то не является ли дополнительная энергетическая стоимость хождения на двух ногах, ценой которой человек получил возможность освободить свои передние конечности для других целей, шагом вперед, давшим ему почти безграничные преимущества перед теми
 |
|
Рис. 31. Чистые затраты кислорода на бег в зависимости от размеров тела у млекопитающих (по Тэйлору и др., 1970). Каждая точка соответствует затрате кислорода на перенос 1 г веса тела на расстояние 1 км (это соответствует наклонам линий на рис. 30). Наклон линии регрессии, наилучшим образом соответствующей полученным точкам, равен —0,40. |
млекопитающими, которые остались четвероногими? Этот вопрос можно изучить (хотя, вероятно, разрешить его и не удастся) на бегающих птицах, которые также передвигаются на двух ногах. Пока что мы изучили только одну из бегающих птиц — южноамериканского нанду, вес которого достигает 20—30 кг. Эта птица, бегающая на двух ногах, также расходует, больше энергии, чем можно предсказать на основании линии регрессии, построенной нами для млекопитающих. А может быть, это характерно и для других птиц?
| {74} |
Все сказанное выше было основано на анализа приращения затрат энергии, вызванного движением; но ведь животное, которое передвигается (например, овца, пасущаяся на лугу), должно, кроме того, непрерывно затрачивать энергию на поддержание своего обычного метаболизма. Давайте проведем следующую аналогию с автомобилем: мы обычно измеряем количество горючего, необходимого для пробега определенного числа километров, не вычитая из этого расходы на поддержание двигателя в рабочем состоянии при нулевой скорости. Можно вычислять приращение затрат энергии на движение или же общий расход энергии в зависимости от задач, которые ставит перед собой исследователь. Например, Тьюкер определял общий расход «горючего» для сравнения затрат на передвижение у ряда животных — летающих насекомых и птиц, шагающих или бегающих млекопитающих,— сопоставляя их с искусственными средствами передвижения, подобно тому как Вейс-Фо (1952) сравнивал насекомых с самолетами. Изящные исследования Тьюкера (1968, б) на летающих птицах дали ценные сведения, расширившие наши знания о локомоции животных. Собранные им данные в совокупности с другими результатами суммированы на рис. 32, который позволяет получить общее представление о затратах энергии, сопряженных с передвижением животных.
Один аспект, касающийся группировки данных по млекопитающим, требует некоторых разъяснений. На рис. 32 данные как для самых мелких, так и для самых крупных млекопитающих лежат гораздо выше линии регрессии, в результате чего создается впечатление, что вся область расположения точек загибается кверху на обоих концах. Черные кружки на рис. 32 идентичны точкам на рис. 31; к ним лишь добавлено несколько точек по литературным данным. Вычисленная линия регрессии для всех этих точек относится к чистой затрате кислорода на движение. Белые же кружки относятся к суммарным затратам кислорода; если вычесть из соответствующих данных потребление кислорода в состоянии покоя, то они сместились бы вниз, приблизившись к линии регрессии. {75}
Этим объясняется кажущееся загибание области расположения точек кверху. Среди исследованных нами млекопитающих у животных средних размеров интенсивность метаболизма во время бега возрастает в 6—8 раз; метаболизм в покое составляет у них, таким образом, весьма незначительную долю суммарного
 |
Рис. 32. Затраты энергии при движении животных, оцениваемые по количеству кислорода, необходимого для перемещения 1 г веса тела на расстояние 1 км. Все данные, за исключением специально оговоренных случаев, относятся к общим метаболическим расходам при движении. Кружки — ходьба и бег. Черные кружки — чистые затраты на движение (те же данные, что и на рис. 31); линия регрессии вычислена по методу наименьших квадратов. Белые кружки — суммарное поглощение кислорода животными при движении, включая поглощение O2 во время отдыха (то есть при нулевой скорости). Поэтому эти точки располагаются выше линии регрессии для чистой затраты. Треугольники — летающие животные. Двойные треугольники, обращенные вершинами друг к другу, — птицы; одинарные — насекомые. Линии регрессии проведены на глаз. Квадраты — плавающие животные (рыбы, за исключением двух квадратов, помеченных «утка» и «человек»); 5 квадратов, лежащих на линии регрессии (проведена на глаз), относятся к нерке (Бретт, 1965); квадрат с флажком — к омулю и хариусу (Матюхин и Столбов, 1970); два светлых квадрата — к лягодону (Вольшлаг и др., 1968); данные для утки — по Пранджу и Шмидту-Нильсену (1970). |
метаболизма, и поэтому вычтем ли мы его величину или нет, практически не играет роли. Среди мелких животных (левый конец) многие не способны к быстрому бегу; поэтому интенсивность их метаболизма во время движения лишь в 2—3 раза выше, чем в покое, то есть у них разница получается {76} довольно существенная. Если вычесть метаболизм в покое из суммарного, то соответствующие точки заметно смещаются вниз. На правом конце лежат точки для лошадей, идущих шагом. У них также метаболизм в покое составляет значительную долю суммарного метаболизма при ходьбе, что приводит к значительному смещению точек вверх. И снова если вычесть метаболизм в покое из суммарного, то эти точки сдвинутся вниз, приблизившись к линии регрессии. Таким образом, весьма вероятно, что загибание области расположения точек для млекопитающих вверх — артефакт, вызванный включением затрат на метаболизм в покое в общие затраты кислорода.
Трудно сказать, какой метод вычисления энергетических затрат на передвижение следует считать наиболее правильным. Если мы хотим выяснить, сколько «горючего» потребляет животное при ходьбе, то следует определять суммарную интенсивность метаболизма. Если же нас интересуют энергетические затраты на локомоцию как таковую, то представляется более правильным установить приращение интенсивности метаболизма в зависимости от приращения скорости. Это приращение остается постоянным для относительно большого диапазона скоростей (см. рис. 30) и позволяет довольно легко вычислить чистую затрату энергии на локомоцию.
Рассмотрим теперь другие способы передвижения. На первый взгляд трудно представить себе, что птица затрачивает на движение меньше энергии, чем наземное млекопитающее такого же размера: ведь ей приходится все время поддерживать вес своего тела в среде, имеющей низкую плотность. Только на то, чтобы удерживать тело в воздухе, птице приходится непрерывно расходовать энергию, не говоря уже о необходимости постоянно преодолевать аэродинамическое сопротивление среды: между тем для бегущего животного возрастание сопротивления воздуха составляет едва заметную долю того сопротивления, которое око преодолевает при передвижении с обычной скоростью. Если, однако, мы вспомним, что во {77} время миграций птица может пролетать без остановки более 1000 км, то окажется очевидным экономическое преимущество полета: едва ли можно представить себе, чтобы мышь сходного веса могла пробежать 1000 км без отдыха и еды.
Если у млекопитающих затраты на бег возрастают в линейной зависимости от скорости, то у птиц положение иное. У волнистых попугайчиков, например, существует некоторая оптимальная скорость полета, при которой потребление кислорода минимально; если
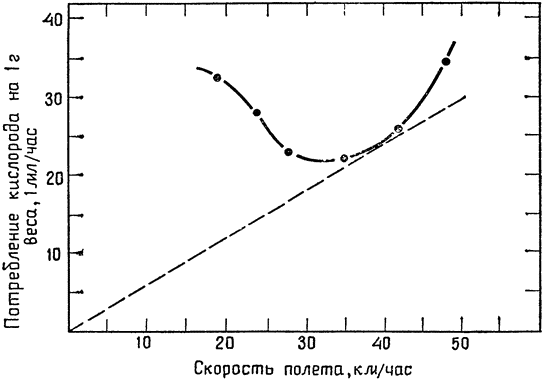 |
Рис. 33. Изменение потребления кислорода у волнистых попугайчиков при разной скорости полета; при скорости 35 км/час потребление кислорода минимальное (по Тьюкеру, 1968, б). |
птица начинает летать быстрее или медленнее, интенсивность метаболизма возрастает (рис. 33). Хотя самое низкое потребление кислорода происходит при полете со скоростью 35 км/час, для того чтобы с наименьшими энергетическими затратами покрыть некоторое расстояние, птица должна лететь несколько быстрее — волнистый попугайчик со скоростью 40 км/час. Это неудивительно; конечно, птица, летящая с большей скоростью, достигает места назначения быстрее. Чтобы установить оптимальную скорость полета на определенное расстояние с наименьшей затратой энергии, достаточно провести из начала координат касательную к кривой (рис. 33); точка касания {78} определяет минимальный наклон этой касательной, то есть минимальные затраты энергии на перемещение.
При изучении полета была обнаружена одна интересная особенность: все данные по насекомым ложатся на линию, служащую продолжением линии регрессии для птиц (см. рис. 32). Некоторые точки для самых мелких насекомых (весом около 1 мг) оказались ниже этой линии, однако все они построены по данным из довольно старых работ. Напротив, новые данные для комаров, весящих около 1 мг, хорошо укладываются на линию регрессии (Нейяр и Ван Хандель, 1971). Поскольку в аэродинамическом отношении между полетом птиц и полетом насекомых имеются существенные различия, возможно, что эта общая линия регрессии носит случайный характер, а не отражает какое-то глубокое сходство между ними.
Как показывает далее рис. 32, энергетические затраты при плавании даже еще ниже, чем при полете. Удельный вес большинства плавающих животных близок к удельному весу воды, поэтому при движении в воде энергия расходуется главным образом на преодоление сопротивления среды, затраты же на поддержание тела в толще воды очень невелики. Поскольку сопротивление среды возрастает пропорционально квадрату скорости, потребность в энергии повышается с увеличением скорости плавания. Сопротивление зависит, кроме того, от площади поверхности животного (а также от других факторов, в том числе от формы); поэтому крупные рыбы благодаря своей относительно меньшей поверхности продвигаются вперед легче и с меньшими усилиями.
Определения энергетических затрат на плавание для нерки различных размеров, начиная с рыбок весом в несколько граммов, показали, что мелким рыбкам плавание «обходится дороже» (Бретт, 1965), Данные по другим лососевым (омулю и хариусу), изучавшимся в других частях света другими авторами (Матюхин и Столбов, 1970), оказались очень сходными. Таким образом, эти определения можно считать достаточно надежными. Весьма близкие результаты были получены при изучении лягодона (Lagodon rhomboides) — рыбы, не относящейся к лососевым {79} (Вольшлаг, Камерон и Цех, 1968). Можно подумать, что многие рыбы отлично знают, во что обходится плавание.
Если сравнивать весьма сходные затраты на плавание у рыб с затратами, производимыми человеком и уткой, плавающими по поверхности воды, то последние оказываются гораздо выше. Плывущая утка расходует в 10 раз больше энергии, чем рыба, — примерно столько, сколько должно расходовать животное
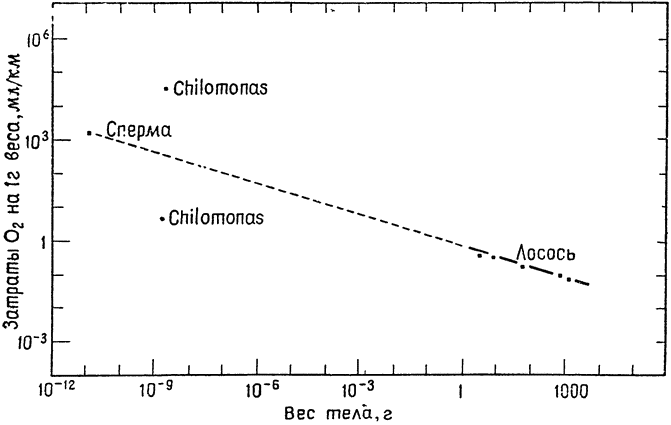 |
|
Рис. 34. Затраты кислорода на плавание у лосося, у спермы быка и у жгутикового простейшего Chilomonas. Chilomonas (вверху) — расчет основан на суммарном потреблении кислорода; Chilomonas (внизу) — расчет основан на вычисленной потребности в энергии для плавания. |
при ходьбе. Человек, организм которого в общем не приспособлен к передвижению в воде, расходует при Плавании в 5 раз больше энергии, чем при беге; это лишнее свидетельство того, что плавание — удел рыб, а полет — удел птиц.
Я вычислил энергетические затраты плавающей в воде спермы на той же основе, на какой определялись затраты энергии у рыб. Используя данные а механической работе, совершаемой при продвижении спермы в воде, а также данные о количестве поглощаемого ею кислорода (Рикменстол, Синтон и Яник, 1969, Ротшильд, 1953); нетрудно вычислить энергию, {80} затрачиваемую 1 г спермы для перемещения на расстояние 1 км. При этом мы исходили из того, что весь поглощаемый спермой кислород затрачивается на плавание, ибо в этом и состоит ее главная деятельность. Результаты оказались очень интересными: точка, вычисленная для спермы быка, точно легла на продолжение линии, характеризующей затраты кислорода при плавании у лосося (рис. 34).
Пытаясь произвести аналогичные вычисления для организма, у которого лишь небольшая и притом неизвестная доля энергии расходуется на плавание, мы сталкиваемся с трудностями. Жгутиковое простейшее Chilomonas, вероятно, расходует весьма малую часть своей энергии на движение вперед. Поэтому если положить в основу расчетов суммарное потребление кислорода, то соответствующая точка оказывается гораздо выше, чем можно было ожидать; если же исходить только из того количества энергии, которое этот организм должен расходовать на движение с обычной для него скоростью по чисто гидродинамическим причинам, то эта точка оказывается гораздо ниже (расчеты основаны на данных Хатченса, Подольского и Моралеса, 1948).
Но вернемся к наземным животным и займемся теперь влиянием активности на количество воды, теряемой ими в результате испарения. Влияет ли учащение дыхания при беге на испарение? Во введении к первой главе мы уже отмечали, что кенгуровая крыса теряет воду главным образом через дыхательные пути. Повышение активности приводит к увеличению интенсивности дыхания, а тем самым и испарения, однако степень этого увеличения предсказать трудно. Испарение через дыхательные пути зависит от температуры выдыхаемого воздуха, которая в свою очередь зависит от теплообмена в носовой полости. Увеличение потока воздуха, проходящего через носовую полость, может понизить эффективность теплообмена, повысить температуру выдыхаемого воздуха, а следовательно, и содержание в нем воды. {81}
Когда кенгуровая крыса неподвижна, ее водный баланс положителен, так как количество воды, образующейся в организме за счет процессов окисления, больше количества воды, испаряющейся через дыхательные пути. Проблема состоит в том, чтобы выяснить, не превышает ли количество воды, испаряющейся из организма в период активности, количества метаболической воды. В качестве меры образования метаболической воды можно использовать потребление кислорода; при этом водный баланс удобно определять как отношение испарения (мг Н2О) к потреблению кислорода (мл О2). Если это отношение ниже 0,6 или 0,7, то водный баланс кенгуровой крысы можно считать положительным, а если выше — то отрицательным. (Точное положение границы между этими двумя режимами зависит от состава пищи и от содержания в ней воды; даже в сухих семенах есть некоторое количество воды.)
Во время бега при умеренной температуре заметных изменений соотношения Н2O : О2 у кенгуровых крыс не наблюдалось (рис. 35). Иными словами, если у животного на отдыхе приход и расход воды сбалансированы, то усиление дыхания при беге не приводит к изменениям водного баланса. Однако при повышении температуры воздуха отношение Н2O : O2 возрастает и водный баланс становится все более неустойчивым. Но, поскольку это отношение при любой данной температуре остается постоянным, независимо от того, отдыхает ли животное или бегает, активность как таковая не оказывает на него непосредственного влияния. Если же испарение усиливается и количество воды, образующейся за счет окислительных процессов, не может покрыть возросшие расходы влаги, то продолжающаяся активность будет увеличивать дефицит воды в организме и животному лучше прекратить движение и укрыться в своей подземной норе. Я не хочу этим сказать, что ночной образ жизни кенгуровой крысы связан со сложностями ее водного баланса; однако то обстоятельство, что ее активность и поиски пищи падают на сравнительно прохладные ночные часы, безусловно благоприятно для водного баланса этого животного. {82}
Что касается животных, активных в дневное время и не имеющих нор, в которых можно укрыться, то для них проблема испарения оказывается гораздо более острой. Для изучения этого вопроса в лабораторных условиях мы выбрали двух животных средних размеров — африканскую гиеновую собаку (Lycaon
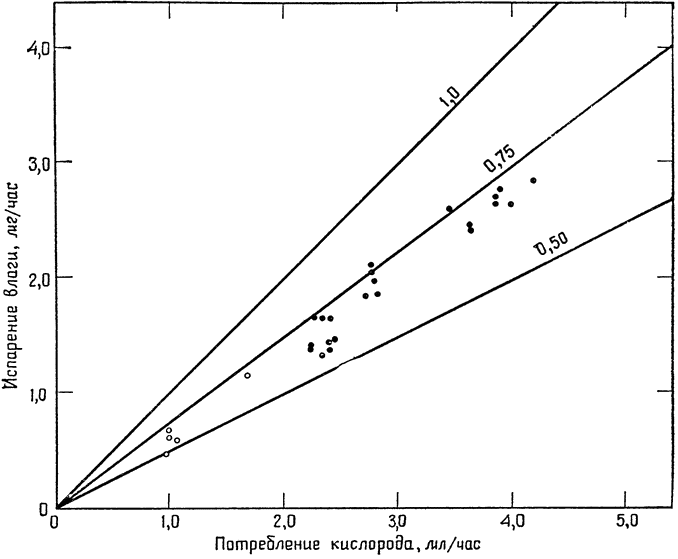 |
|
Рис. 35. Зависимость испарения влаги через дыхательные пути от поглощения кислорода у кенгуровой крысы (по Раабу и Шмидту-Нильсену, 1971). Температура среды 25°С. Белые кружки — во время отдыха; черные кружки — при беге. Цифры на прямых указывают величину их наклона. |
pictus) и крупную птицу — южноамериканского нанду. Оба эти вида — хорошие бегуны, и их можно научить бегать по тредбану.
Много лет назад мы установили, что верблюд знойным днем в пустыне уменьшает потери воды, допуская повышение температуры тела, вместо тог» чтобы расходовать воду на охлаждение (Шмидт-Нильсен и др., 1957). Конечно, такое повышение температуры тела ограничено: максимальное повышение, {83} которое нам приходилось наблюдать, составляло 7°С — от 34°С утром температура повысилась до 41°С в жаркий полдень. Высокая температура тела создает ряд возможностей для экономии воды. Прежде всего она позволяет избежать расхода воды на рассеивание тепла, идущего на повышение температуры тела, а ночью, когда становится прохладнее, это избыточное тепло можно отдать среде опять-таки без затрат воды. Затем, что гораздо важнее, при высокой температуре тела перепад температур между телом животного и очень жаркой окружающей средой может уменьшиться и даже изменить знак; тем самым животное получает возможность терять тепло, не расходуя воды. А нельзя ли предположить, что сильное повышение теплопродукции у животного во время бега также создает ему известное преимущество?
Гиеновая собака — известный хищник, который охотится днем, то есть во время жары. Она берет не быстротой бега, а главным образом выносливостью, неутомимо преследуя свою жертву. Чтобы носиться по жарким равнинам Африки, поддерживая постоянную невысокую температуру тела, ей пришлось бы интенсивно испарять воду, а это, по всей вероятности, сильно ограничивало бы расстояние, которое она могла бы пробегать. Высокая же температура тела позволила бы гиеновой собаке сэкономить воду.
Гиеновая собака — довольно неподатливое животное, которое не удается приручить. При содержания в неволе самки неизменно убивают своих щенят, и, чтобы вырастить животных, их сразу отнимают от матери и выкармливают искусственно. Получив новорожденного щенка из вашингтонского Национального зоопарка, мы быстро поняли, почему никто не стремится одомашнивать гиеновую собаку — один только запах уже служит достаточным препятствием. Смирившись с некоторыми неприятными обонятельными ощущениями, мы научили вскормленное нами животное бегать по тредбану с надетой на морду маской, которая позволяла регистрировать потребление кислорода. Наблюдения за тепловым балансом гиеновой собаки показали, что у нее действительно достигается значительная экономия воды за счет повышения температуры тела (Тэйлор и др., 1971, б). Во время отдыха {84} при комнатной температуре ее тело примерно на один градус теплее, чем у домашней собаки, а при ускорении бега температура у гиеновой собаки повышалась быстрее, чем у домашней (рис. 36). При достижении скорости 15 км/час температура тела домашней собаки повысилась на 1,3°С, а гиеновой — на
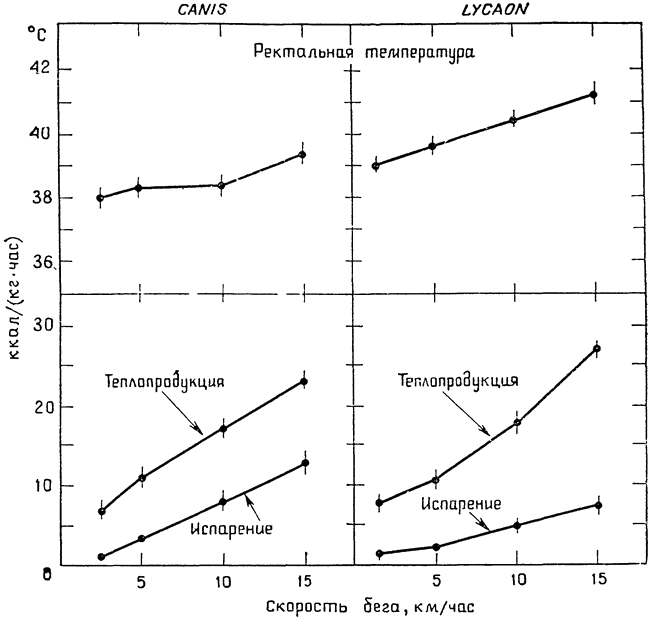 |
|
Рис. 36. Сравнение теплового баланса домашней (Canis) и гиеновой (Lycaon) собак (по Тэйлору и др., 1971, б). Во время бега температура тела у Lycaon достигает более высокого уровня, чем у Canis (верхние графики); этому соответствует относительно слабое испарение воды через дыхательные пути (нижние графики). Эксперименты проводились при 26°С и 25%-ной относительной влажности. |
2,2°С и стала равной 41,2°С. Оба животных бегали при комнатной температуре; чем же отличался их тепловой баланс?
Во время бега теплопродукция (определяемая по потреблению кислорода) у гиеновой собаки была несколько выше, чем у домашней; но если обратить внимание на использование воды (нижние графики {85} на рис. 36), то окажется, что у гиеновой собаки при беге со скоростью 15 км/час только 25% производимого тепла рассеивалось через дыхательные пути (тепловая одышка), тогда как у домашней собаки таким путем рассеивалось до 66% тепла. Вывод напрашивается сам собой: гиеновая собака действительно сокращает расход воды, допуская -повышение температуры тела, что позволяет организму отдавать тепло за счет теплопроводности. Если потери воды уменьшаются вдвое, то расстояние, в пределах которого этот хищник может преследовать свою жертву, очевидно, удваивается (при допущении, что лимитирующим фактором для этого расстояния является вода).
Я уже неоднократно подчеркивал, что повышение температуры тела при тепловых нагрузках нельзя считать показателем «несрабатывания» терморегуляции (Шмидт-Нильсен, 1964). А теперь мы видим, что и повышение температуры животного при беге можно истолковать как способ уменьшения потерь воды, а не как проявление недостаточного рассеивания тепла.
Давно известно, что у человека при физической нагрузке температура тела повышается и притом в строгой зависимости от интенсивности этой нагрузки. Если нагрузка поддерживается на постоянном уровне, то ректальная температура остается несколько повышенной, причем ее уровень определяется интенсивностью нагрузки (Нильсен и Нильсен, 1962). Обычно это объясняют тем, что повышенная температура тела ускоряет все физиологические процессы и тем самым способствует выполнению физических упражнений. По моему мнению, для организма, эволюция которого была направлена на развитие способности преследовать добычу, пробегая большие расстояния по открытой местности, возможность экономить воду благодаря повышенной температуре тела следует считать отнюдь не менее важным преимуществом.
Рассмотрим теперь нанду. Это крупная птица весом около 25 кг, хотя и поменьше африканского страуса, вес которого может достигать 100 кг. В естественных {86} условиях нанду бегает довольно быстро, но в отличие от человека и собаки он скорее спринтер, чем стайер. Поэтому интересно было выяснить, каким образом у этой птицы происходит терморегуляция. Приручить нанду бегать в течение длительного времени с равномерной скоростью оказалось трудным делом; их удавалось заставить бегать при высокой температуре лишь очень недолго. Поэтому была выработана следующая процедура эксперимента: птицу
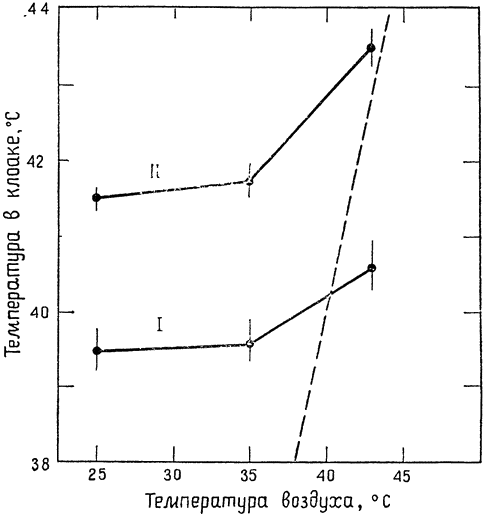 |
Рис. 37. Повышение температуры тела у нанду после бега (по Тэйлору и др., 1971, а). I — температура тела до бега; II — температура тела после бега. Бег со скоростью 5 км/час в течение 20 минут вызвал повышение температуры тела примерно на два градуса. |
заставляли бежать в течение 20 минут, а затем давали ей 40 минут отдыха; на протяжении всего этого часа регистрировались продукция метаболического тепла и рассеивание тепла.
Бег в течение 20 минут всегда приводил к значительному повышению температуры тела нанду: при комнатной температуре — на 2°С, а при 43°С — немного более чем на 2,5°С (рис. 37). {87}
Поскольку температура тела нанду к концу бега при температуре воздуха 43°С несколько выше последней, его тело может отдавать среде некоторое количество тепла за счет теплопроводности, и излучения; тем самым количество тепла, рассеиваемого путем испарения, уменьшается. Если же птица отдыхает при 43°С, то температура тела оказывается ниже температуры среды, тепло поступает в ее тело за счет
 |
|
Рис. 38. Тепловой баланс у нанду (по Тэйлору и др., 1971, а). А. Отдых в течение часа. Б. 20-минутный бег с последующим 40-минутным отдыхом. I — теплопродукция; II — суммарное испарение; III — испарение из дыхательных путей. Разность между теплопродукцией (I) и суммарной потерей за счет испарения (II) равна теплоотдаче за счет теплопроводности и излучения (заштрихованная область). Отдыхающая птица (A) получает некоторое количество тепла из среды (область, покрытая точками). После бега (Б) температура тела птицы повышается и перепад температур между ее телом и средой дает ей возможность продолжать теплоотдачу за счет теплопроводности и излучения (заштрихованная область). |
теплопроводности и это избыточное тепло ей приходится рассеивать путем испарения вместе с метаболическим теплом. Как же влияет изменение температуры тела на использование воды?
Бег со скоростью 5 км/час (для нанду такую скорость можно считать умеренной) вызывает повышение теплопродукции за счет метаболизма в 2,5 раза независимо от температуры среды. В течение часа, слагающегося из 20-минутного бега и следующего за ним 40-минутного отдыха, испарение у нанду закономерно возрастало с повышением температуры воздуха (рис. 38, Б). Если же птица в течение всего часа отдыхала, испарение также увеличивалось в зависимости от температуры, но в меньшей степени (рис. 38, А). При сравнений теплопродукции и потерь тепла за счет одних только теплопроводности и конвекции, без испарения; у активного и отдыхающего животного оказывается, что в первом случае они гораздо — почти втрое — больше, причем главным образом за счет повышения температуры тела.
Птицы, бегавшие при высоких температурах воздуха, не выходили на стационарный режим. Они не могли или не хотели продолжать бег, температура их тела не стабилизировалась, и, по всей вероятности, они оказались бы неспособными к длительному бегу в очень жаркой среде. Вопросы о том, какой степени перегрева достигли бы нанду, если бы их заставили продолжать бег при высокой температуре, и сколь высокие температуры тела они могут выдержать, остаются пока без ответов. Возможно, эти птицы и переносили бы очень высокие температуры тела, если бы они могли «держать головы в холоде»; к рассмотрению этого вопроса мы перейдем в следующей главе.
| {89} |
Недавно Тэйлор установил, что у газелей и антилоп температура тела может подняться выше 46°С, причем это, по-видимому, не причиняет им никакого вреда. Бейза (Oryx beisa), вес которой достигает 100 кг, очень хорошо переносит сухую жаркую погоду и может простоять на ярком солнце целый день. Если содержать это животное на сухом корме, не давая воды, то потребление кислорода у него значительно уменьшится, что приводит к снижению внутренней тепловой нагрузки. Хотя при очень высокой температуре воздуха бейза испаряет воду за счет тепловой одышки, количество расходуемой воды резко сокращается, поскольку температура ее тела повышается при этом до более чем 45°С. В опытах Тэйлора бейза выдерживала повышение ректальной температуры до 46,5°С в течение 6 часов без каких-либо заметных вредных последствий.
Каким образом животное, принадлежащее к млекопитающим, может выдерживать такую высокую температуру тела? Наиболее чувствительные к повышению температуры части организма — это центральная нервная система, и способность выдерживать температуру 46°С следовало бы считать совершенно исключительным для высших животных явлением (Тэйлор, 1970). Ответ оказался простым: температура головного мозга не повышается так сильно, как температура тела. У бейзы, температура тела которой повышалась под влиянием физической нагрузки, температура головного мозга оказалась на 2,9°С ниже температуры крови в артериях, находящихся в глубоких областях тела (Тэйлор, 1969). Кровь, направляющаяся к мозгу, охлаждается благодаря особому расположению сосудов в голове. Наружные сонные {90} артерий, по которым в основном происходит кровоснабжение головного мозга, проходят через кавернозный синус, где они распадаются на сотни мелких артерий. Кавернозный синус наполняется венозной кровью, оттекающей из сосудов носовой полости, где она охлаждается благодаря испарению с влажных слизистых. Артериальная кровь на пути к мозгу протекает
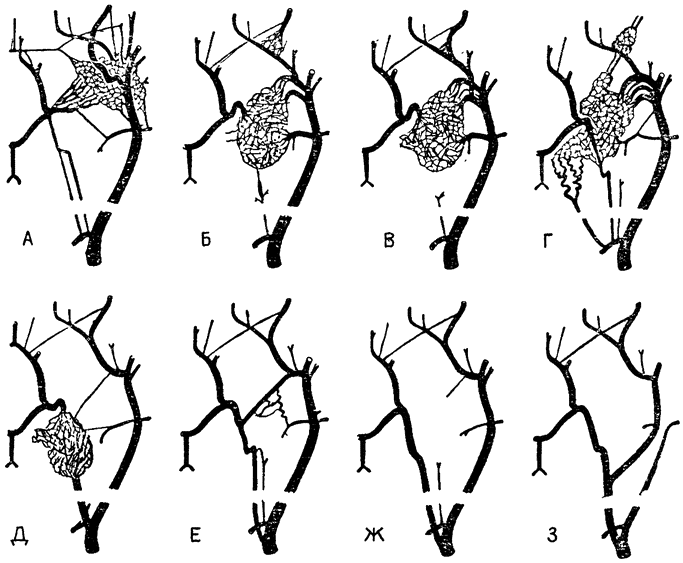 |
|
Рис. 39. Каротидная сеть у разных животных (по Даниэлю, Доуэсу и Причарду, 1953). А. Домашняя кошка (Felis domestica). Б. Баран (Ovis aries). В. Безоаровый козел (Capra hircus). Г. Бык (Bos taurus). D. Кабан (Sus scrofa). E. Домашняя собака (Canis familiaris). Ж. Европейский кролик (Oryctolagus cuniculus). З. Пасюк (Rattus norvegicus). |
по сосудам, проходящим через это скопление охлажденной крови, и ее температура неизбежно понижается. Роль каротидной сети в создании разницы между температурой тела и температурой крови, снабжающей мозг, показана на ряде животных Бейкером и Хэйуордом (1968). В результате тщательных измерений, проведенных на овцах, кровообращение которых сходно с кровообращением у других парнокопытных, {91} они установили, что венозная кровь, возвращающаяся из слизистой носовой полости в кавернозный синус, действительно охлаждает артериальную кровь в каротидной сети, а это имеет важное значение для поддержания на допустимом уровне температуры гипоталамуса у овцы во время тепловой одышки. Аналогичные сетчатые образования, охлаждаемые венозной кровью, возвращающейся из области носа, имеются у многих животных, хотя и не у всех (рис. 39). У собаки эта сеть устроена сложнее; она образует анастомозы между наружными и внутренними сонными артериями, создавая возможность, по терминологии Бейкера и Хэйуорда, для «двух типов регуляции температуры мозга». У крысы нет сетчатых или сходных с ними структур, однако ей это и не нужно, поскольку она не прибегает к тепловой одышке, а ее носовая полость, как уже отмечалось в главе 1, приспособлена к удержанию, а не к потере тепла.
Создается впечатление, что теплообмен в каротидной сети дает животным, прибегающим к тепловой одышке, важное преимущество по сравнению с животными, охлаждающими свое тело путем потоотделения. Такой теплообменник позволяет поддерживать низкую температуру в какой-то одной части тела, в данном случае — в мозгу, что, очевидно, снижает общее количество расходуемой воды. А при терморегуляции с помощью потоотделения животное, по-видимому, должно охлаждать весь свой организм до той температуры, превышение которой вредно для головного мозга, что требует гораздо больших затрат воды1.
| {92} |
Теплообменник, создаваемый каротидной сетью, в принципе аналогичен теплообменнику в плавниках кита (см. стр. 16). В обоих случаях температура какой-то части организма (в одном — головного мозга, а в другом — плавника) поддерживается на более низком уровне, чем температура всех остальных частей. Разница заключается в том, что у кита низкая температура плавника позволяет сохранить тепло глубоких участков тела, а антилопа «держит голову
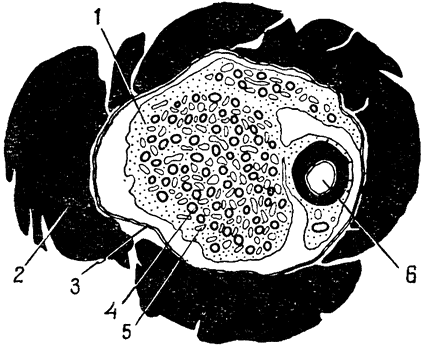 |
|
Рис. 40. Кровоснабжение семенника крупного сумчатого — валлаби (по Барнетту, Харрисоку и Томлинсону, 1958). 1 — соединительная ткань; 2 — мышцы; 3 — оболочка семенника; 4 — артерия; 5 — вена; 6 — семяпровод. |
в холоде», чтобы иметь возможность поднять температуру остальных частей тела выше уровня, допустимого для мозга.
Аналогичный теплообмен происходит также в сосудистых пучках, которые снабжают кровью, например семенники барана, и особенно хорошо развиты у некоторых крупных сумчатых (рис. 40). Благодаря относительно слабому испарению с поверхности мошонки температура семенников может поддерживаться на более низком уровне, чем температура организма в целом, а сосудистый теплообменник предотвращает «растворение этого холода» в общей массе тела. Испарение с поверхности мошонки могло бы вызвать лишь очень незначительное охлаждение глубоких {93} участков тела, так что если бы не было теплообменника, то семенники снабжались бы кровью, имеющей температуру тела, а это по малопонятным причинам препятствует образованию нормальной жизнеспособной спермы.
Независимая от остального тела регуляция температуры в отдельных частях организма создает также высокую температуру в мышцах некоторых крупных быстроплавающих рыб, таких, как тунцы и акулы. У большинства рыб температура тела очень близка к температуре окружающей воды. Кровь, которая поглощает из воды кислород, протекает через жабры, где она охлаждается до температуры воды; поэтому температура тела рыбы может повыситься только в том случае, если между ее жабрами и остальными тканями имеется теплообменник. У тунца кровеносные сосуды, снабжающие красные боковые мышцы1 туловища, образуют слой тесно переплетающихся между собой мелких артерий и вен, по которым кровь течет в противоположных направлениях (рис. 41).
В этот теплообменник поступает артериальная кровь, приходящая от жабр; она течет по сосудам, лежащим близко к поверхности тела рыбы, и поэтому ее температура соответствует температуре воды. Таким образом, холодный конец теплообменника находится на поверхности рыбы, а теплый — внутри, в толще мышц. Благодаря этому теплообменнику у тунца температура мышц может быть на целых 14°С выше температуры воды, в которой она плавает (Карей и Тиль, 1966). Сходная система имеется у серо-голубой акулы (Jsurus oxyrhynchus) и, по всей вероятности, у некоторых других видов акул, у которых обнаружены подобные сосудистые пучки (Карей и Тиль, 1969). Такая «теплокровность», вероятно, дает некоторые преимущества этим крупным быстроплавающим рыбам.
| {94} |
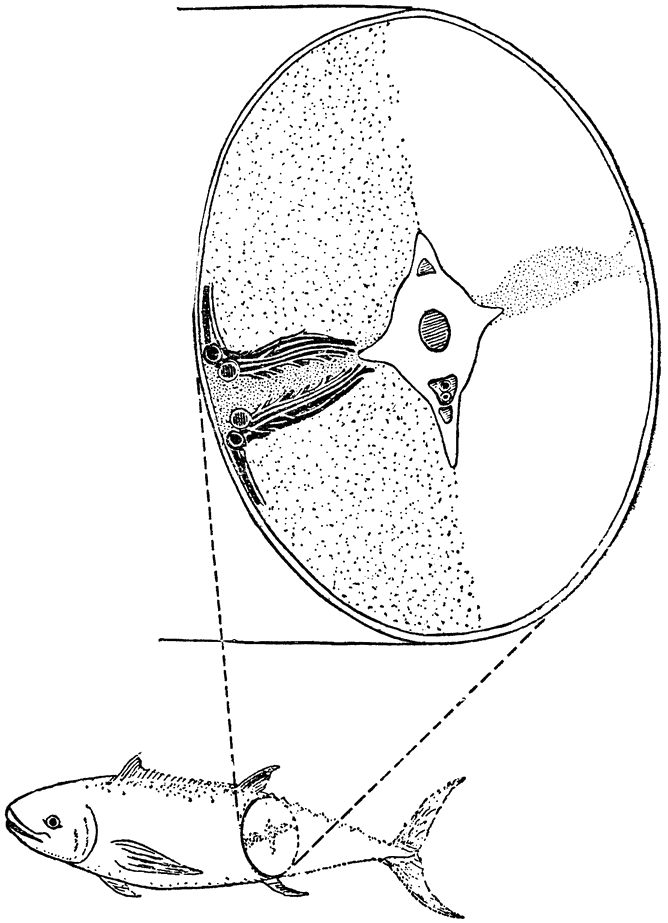 |
|
Рис. 41. Теплообменник в системе кровоснабжения красных мышц, поддерживающий их температуру на более высоком уровне (по Карею и Тилю, 1966). Мелкие артерии, берущие начало от более крупной кожной артерии, образуют слой, в котором они тесно переплетаются с венами. |
| {95} |
Есть один вопрос, которого мы до сих пор не касались. Как уже говорилось, в плавнике кита тепло передается от артериальной крови венозной, никогда не доходя до поверхности плавника. Если бы кислород передавался с такой же легкостью, то эффективный теплообменник препятствовал бы снабжению кислородом периферических тканей. Вероятно, именно беспрепятственной диффузией кислорода между капиллярами, приносящими кровь в почечные сосочки, и капиллярами, уносящими кровь (нисходящая и восходящая ветви vasa recta), объясняется очень низкое содержание кислорода в почечной ткани, столь обильно снабжаемой кровью (Ландис, Леонгардт и Деремен, 1964).
Однако, исследуя теплообменники животных, мы обнаружили, что диаметр образующих их кровеносных сосудов равен примерно 1 мм. Как известно, кислород очень слабо диффундирует через стенки сосудов такого размера, так что эти теплообменники плохо приспособлены для обмена кислорода. Это можно было бы выразить количественно, сравнивая коэффициенты диффузии для тепла и кислорода, если бы нам удалось найти подходящий физиологический эквивалент для этих двух потоков крови, проходящих через теплообменник.
Кислородная емкость млекопитающих равна примерно 200 мл кислорода на 1 л крови. Содержащийся в 1 л крови кислород при полном использовании (физиологически допустимый максимум) дает 1000 калорий тепла; иными словами, он может повысить температуру одного литра крови на один градус, но не более того. Таким образом, мы установили своего рода физиологический эквивалент для сопоставления кислорода и температуры: 150-миллиметровый градиент содержания кислорода (различие между полностью насыщенной кислородом кровью и нулевым его напряжением в тканях) может создать температурный градиент в 1°С, так как в стабильном состоянии каждый литр крови может принести тканям упомянутое выше количество кислорода и забрать у них эквивалентное количество тепла. Если кислородная емкость {96} крови ниже (например, у рыб она вдвое ниже, чем у млекопитающих), то максимальный перепад температур соответственно меньше, однако в принципе картина остается прежней.
Теперь мы, пожалуй, готовы к тому, чтобы выяснить, действительно ли диффузия кислорода в теплообменнике так велика, что плавник кита или, что еще хуже, семенники кенгуру получают мало кислорода или не получают его вовсе. Если эффективность теплообменника составляет 99%, то это означает, что 99% тепла, приносимого с артериальной кровью, возвращается венозной крови и только 1% попадает на периферический конец теплообменника; Как показывает простой расчет, теплообменник, допускающий перенос 99% тепла, которое потенциально может дать содержащийся в артериальной крови кислород, допустит диффузию в венозную кровь лишь 2% этого кислорода (которые тем самым не смогут дойти до периферического конца теплообменника и до дистально расположенных тканей).
Для теплообменника 99%-ная эффективность более чем достаточна. Потеря артериальной кровью 1% тепла, например в плавнике кита, не имеет для животного практически никакого значения. Столь же незначительно и то обстоятельство, что 2% кислорода артериальной крови возвращается в вены, не доходя до периферии. Если бы эффективность теплообменника была ниже, то количество «теряемого» артериальной кровью кислорода было бы меньше; ведь, поскольку коэффициенты диффузии остаются неизменными, количества тепла и количества кислорода должны меняться в одинаковой пропорции. Таким образом, теплообменник с эффективностью лишь 95%, который допускает потерю 5% тепла из артериальной крови, допустит диффузию в венозную кровь не более 0,4% кислорода.
Если теперь мы вернемся к противоточным механизмам, связанным с диффузией растворенных веществ, а не тепла, то обнаружим, что их размеры близки к размерам капилляров — примерно 0,01 мм в диаметре. Меньшие размеры сосудов дают двоякий эффект: 1) общая площадь стенок увеличивается, поскольку число сосудов значительно выше; 2) стенка, {97} через которую происходит диффузия, гораздо тоньше. В чудесной сети плавательного пузыря рыб, который мы подробно рассмотрим несколько ниже, обеспечена эффективная диффузия газа между тысячами плотно упакованных тонкостенных капилляров. В легком птицы при диффузии газов между кровью и воздухом должно преодолеваться расстояние около 1 мкм, что не препятствует быстрой диффузии. В носовой полости мелких млекопитающих перемещение тепла и водяных паров происходит с большой скоростью; конечно, размеры этой полости гораздо больше, чем ширина капилляров, однако диффузия водяных паров в воздухе происходит примерно в 100 000 раз быстрее, чем диффузия какого-либо газа в тканях.
Итак, противоточные системы, используемые для теплообмена, состоят из сосудов миллиметрового размера и поэтому не могут функционировать как газообменники. Относительно медленная диффузия газов в теплообменнике создает известное преимущество, так как в противном случае они не имели бы столь широкого распространения. В эффективно действующем газообменнике сосуды должны иметь размеры, близкие к размерам капилляров.
Все тепло- и газообменники, которые мы до сих пор рассматривали, основаны на нисходящих градиентах: от теплого — к холодному, от высокой концентрации газа — к низкой. Между тем существуют структуры, способные действовать «вверх», то есть вызывать перемещение вещества против его градиента концентрации; эти структуры по своему строению и даже свойствам несколько сходны с обменниками, что нередко приводит к довольно сильной путанице.
Структуры эти известны под названием умножителей, или, точнее, противоточных умножителей. Например, в почках млекопитающих происходит активный перенос ионов натрия, что в конечном итоге приводит к образованию концентрированной мочи; поскольку умеренный перенос ионов постепенно создает высокую концентрацию, весь механизм получил {98} название умножителя. В плавательном пузыре рыбы происходит выделение газа, преодолевающее давление в несколько сотен атмосфер, опять-таки при помощи умножителя. Необходимо четко разъяснить, чем и как умножители отличаются от обменников; тогда черты сходства между ними перестанут создавать путаницу.
Почка. Низшие позвоночные неспособны вырабатывать мочу, осмотическое давление которой было бы выше, чем у плазмы крови; такой способностью обладают только птицы и млекопитающие. Механизм, при помощи которого это достигается, описан в большинстве современных учебников физиологии; поэтому здесь мы рассмотрим его лишь в общих чертах, не вдаваясь в подробности.
В почке млекопитающих каждый почечный каналец образует петлю в форме шпильки, которая называется петлей Генле. Одна из ее ветвей, ведущая в почечный сосочек, называется нисходящей, другая — восходящей ветвью. Ионы натрия извлекаются из жидкости, находящейся в восходящей ветви, в результате процесса активного переноса; «активным» он называется потому, что при этом ионы из области более низкой концентрации переносятся в область более высокой концентрации и это требует затраты энергии. Активный перенос приводит к повышению концентрации натрия в окружающих тканях, так что ионы натрия могут поступать в нисходящую ветвь путем простой диффузии. Поскольку из жидкости, покидающей петлю Генле, непрерывно удаляется натрий, она содержит его меньше, чем жидкость, поступающая в эту петлю. Концентрация натрия в перегибе петли Генле становится все выше и выше. Поэтому концентрация натрия в сосочках может достигнуть очень высокого уровня, что делает возможным извлечение воды из жидкости, протекающей по собирательным трубочкам, за счет осмоса (рис. 42).
По причинам, которые нам нет нужды разбирать подробно, максимальная концентрация, достигаемая в этом почечном умножителе, зависит от длины петли Генле. Следует подчеркнуть, что по двум ветвям этой петли жидкость течет в противоположных направлениях и что функция противоточного умножителя
| {99} |
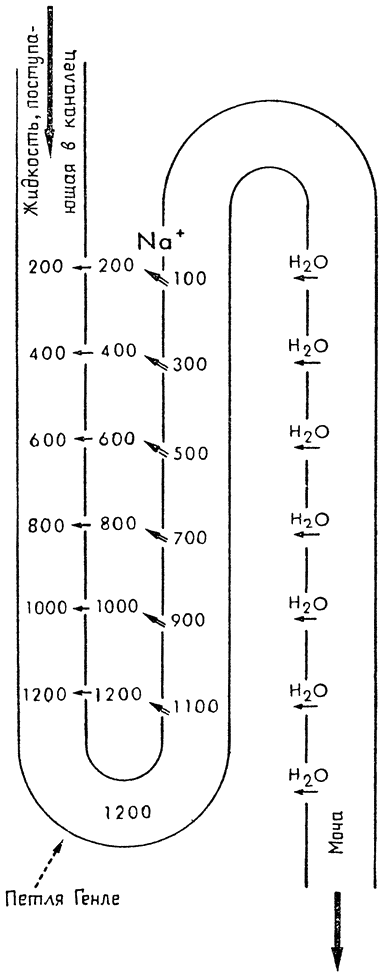 |
|
Рис. 42. Роль петли Генле в качестве противоточиого умножителя в почке млекопитающих. Цифры указывают концентрации натрия; они выбраны произвольно, чтобы показать повышение концентрации в направлении перегиба петли. Маленькими косыми стрелками показан активный перенос ионов натрия, а горизонтальными стрелками — их пассивная диффузия. Натрий в результате активного переноса извлекается из восходящей ветви и путем диффузии проникает в нисходящую ветвь, накапливаясь таким образом в перегибе петли Генле и в интерстициальной ткани. Затем вода осмотически извлекается из собирательных трубочек, в которых остается концентрированная моча. |
| {100} |
целиком зависит от двух предпосылок: 1) наличия активного переноса, осуществляемого с затратой энергии, и 2) протекания жидкости по петле, имеющей форму шпильки, что делает возможным эффект умножения.
Плавательный пузырь. Рыба, живущая на глубине, скажем, 1000 м, испытывает давление со стороны окружающей ее воды, в данном случае равное 100 атмосферам. Газ, наполняющий эластичный плавательный пузырь, испытывает такое же давление (и такое же давление испытывал бы наполненный воздухом резиновый баллон, погруженный в воду на такую же глубину). Однако в окружающей рыбу воде парциальное давление растворенных газов равно лишь 1 атмосфере, так как количество газов, содержащихся в любом данном объеме воды, пришедшем в равновесие с атмосферным воздухом на поверхности, не изменится, если переместить этот объем на глубину 1000 м.
Таким образом, рыба сталкивается с двумя проблемами. Во-первых, для того чтобы плавательный пузырь мог наполниться, парциальное давление газов в нем должно стать значительно выше, чем 1 атмосфера (в рассматриваемом нами случае оно должно увеличиться в 100 раз), а это, очевидно, может произойти только путем активного переноса, то есть с затратой энергии. Во-вторых, как только газ в плавательном пузыре окажется под высоким давлением, необходимо воспрепятствовать его диффузии в кровь, которая в жабрах приходит в состояние диффузионного равновесия с низким парциальным давлением газа в окружающей воде.
Как всегда, для того чтобы понять ту или иную функцию, нам надо сначала получить некоторое представление о структуре выполняющего ее органа. В стенке плавательного пузыря большинства рыб имеется специальная газовая железа, которая снабжается кровью через своеобразную структуру, называемую «чудесной сетью» (rete mirabile). Форма этой сети и самой газовой железы весьма разнообразна у разных видов, однако общий принцип ее устройства всегда один и тот же: артерия, снабжающая кровью газовую железу, распадается на бесчисленное множество прямых, параллельных друг другу капилляров, которые {101} затем вновь собираются в один сосуд, идущий к железе. Вена, уносящая кровь от газовой железы, точно так же распадается на множество параллельных капилляров, которые с поразительной упорядоченностью распределяются между артериальными капиллярами, а затем вновь сливаются вместе, образуя одну нормальную вену.
Капилляры чудесной сети достигают довольно значительной длины — до 25 мм у некоторых глубоководных рыб. Для капилляров такая длина чрезвычайно велика: вообще самыми длинными считаются капилляры мышц, достигающие всего 0,5 мм. У тех рыб,
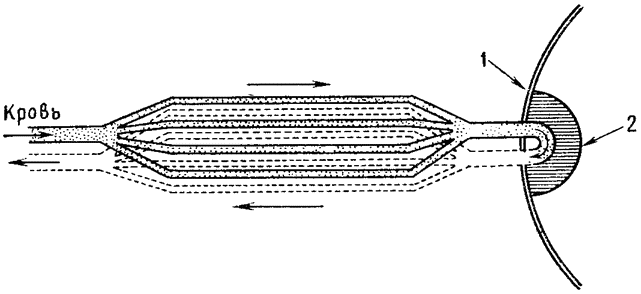 |
|
Рис. 43. Схема чудесной сети в системе кровообращения плавательного пузыря. 1 — стенка плавательного пузыря; 2 — газовая железа. |
которым приходится секретировать газ при наиболее высоких давлениях, капилляры, по-видимому, имеют наибольшую длину; этого и следовало ожидать, поскольку эффективность умножителя зависит от его длины. На рис. 43 схематически изображена чудесная сеть плавательного пузыря. Следует сказать несколько слов о количественном аспекте ее строения. Крог (1929) исследовал с этой точки зрения артериальную и венозную сети угря весом в несколько сотен граммов. Эти сети образованы капиллярами длиной по 4 мм и общим объемом 64 мм3 — примерный объем капли воды. Он насчитал свыше 100 000 артериальных капилляров и примерно столько же венозных, что составляет по 400 м капилляров каждого типа. Их диаметр, как и у большинства других капилляров, равен {102} 7—10 мкм; простой арифметический расчет показывает, что суммарная площадь поверхности стенок капилляров каждого типа составляет по 100 см2 — и все это помещается в объеме, соответствующем объему капли воды! А для нас самое важное заключается в том, что через всю эту огромную поверхность может происходить диффузия.
Рассмотрим сначала, каким образом удерживается в плавательном пузыре газ, выделенный в него газовой железой; процессом секреции газа мы займемся позднее. Здесь наблюдается совершенно то же самое, что и в плавнике кита. Кровь, оттекающая от плавательного пузыря, насыщена газами, которые растворялись в ней при высоком давлении, но, протекая по чудесной сети, она встречается с артериальной кровью, в которой парциальное давление газов ниже. Благодаря большой поверхности, через которую может происходить диффузия, избыточный газ диффундирует в эту артериальную кровь, и в вытекающей из чудесной сети венозной крови парциальное давление газа снижается до его уровня в притекающей артериальной крови. Несмотря на высокое давление газа в плавательном пузырей низкое в артериальной крови, газ остается в пузыре; чудесная сеть служит шлюзом или шлагбаумом, через который газ пройти не может.
Обращаясь к проблеме наполнения плавательного пузыря газом, проще всего начать с кислорода, тай как чаще всего в пузырь секретируется именно этот газ. На рис. 44 чудесная сеть сведена к одной-единственной петле, что позволяет значительно упростить схему. Допустим, что процесс секреции начинается с выделения газовой железой молочной кислоты (которую она и выделяет в действительности). Эта молочная кислота поступает в кровь, снижая ее сродство к кислороду. Вообще действие, оказываемое кислотой на кровь, известно под названием эффекта Бора; однако у большинства рыб это действие выражено гораздо сильнее, чем у млекопитающих, и его называют эффектом Роота (Шоландер и Ван Дам, 1954). Под действием кислоты гемоглобин отдает кислород, и содержание растворенного (то есть не связанного с гемоглобином) кислорода в оттекающей из газовой железы венозной крови повышается. Поэтому {103} кислород диффундирует через артериальные капилляры, где концентрация его ниже, и накапливается в петле. До тех пор, пока содержание кислорода в оттекающей венозной крови ниже, чем в притекающей артериальной, он будет накапливаться все в большем количестве. Благодаря противоточной системе молочная кислота также задерживается в колене петли, усиливая эффект. Конечно, для того чтобы кислород мог накапливаться в петле, его общее количество в венозной крови, вытекающей из сети, должно быть меньше, чем в притекающей артериальной крови; но
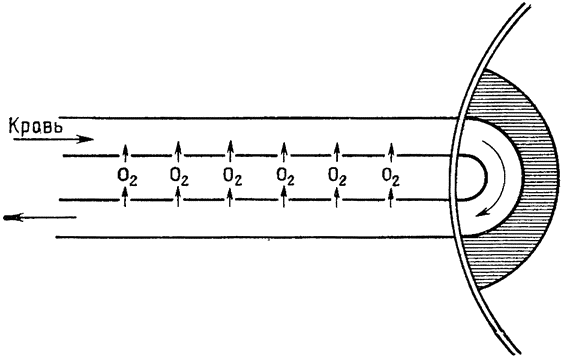 |
Рис. 44. Схема кровообращения в чудесной сети и газовой железе, сведенная к одной петле. |
при этом для того, чтобы кислород диффундировал из венозных капилляров в артериальные, его парциальное давление в венозных капиллярах должно быть выше.
Норвежский ученый Стеен выяснил, что один из факторов, играющих важную роль в создании такого положения вещей, имеет отношение к кинетике реакций связывания гемоглобина с кислородом. По мере поступления в кровь молочной кислоты гемоглобин отдает кислород; эта реакция протекает очень быстро, с полупериодом 50 мс. Реакция, идущая в противоположном направлении, — связывание кислорода гемоглобином — напротив, протекает медленно, с полупериодом 10—20 с (Берг и Стеен, 1967). Это означает, {104} что кислород, освободившись из гемоглобина в петле, не имеет времени соединиться с гемоглобином за то недолгое время, пока кровь течет по венозным капиллярам, и уходит из них; поэтому парциальное давление кислорода в венозных капиллярах остается высоким и он может диффундировать в артериальные капилляры и накапливаться в петле.
В результате такой постоянной диффузии кислорода в артериальные капилляры содержание постепенно накапливающегося в петле кислорода может достигнуть очень высокого уровня. Функция самой газовой железы подробно не изучена, однако теперь нам по крайней мере известно, каким образом в крови, поступающей в железу, создается очень высокое парциальное давление кислорода.
Помимо молочной кислоты повышение парциального давления кислорода может вызываться присутствием других растворенных веществ; растворимость кислорода (и других газов в растворе) понижается в результате эффекта «высаливания». Поскольку в плавательном пузыре может происходить также секреция азота, причем у некоторых рыб секретируется главным образом именно азот, то эффект высаливания, возможно, оказывает в чудесной сети примерно такое же действие, как было описано выше для кислорода (Кюн и др., 1963). Это очень полезно, потому что, хотя мы не можем рассматривать действие, оказываемое кислотой на гемоглобин, как способ накопления инертных газов, мы все же имеем основание считать чудесную сеть противоточным умножителем, позволяющим на основе очень слабого эффекта достигнуть высокого парциального давления газа.
Подводя итоги, можно констатировать, что противоточный умножитель, обусловливающий секрецию газа под высоким давлением в плавательный пузырь, обладает двумя необходимыми умножителю качествами: 1) секреция происходит против градиента концентрации, то есть в «восходящем» направлении; 2) эффект умножения становится возможным благодаря наличию структуры в форме петли или шпильки для волос.
| {105} |
Не все сосудистые сети могут функционировать как противоточные обменники. Для обменника необходимо прежде всего наличие двух потоков. У многих китов и дельфинов имеются обширные густые сплетения сосудов, связанные с артериями, которые направляются к черепу и идут вдоль части позвоночника. Но, поскольку артерии не перемежаются с аналогичными венозными сетями, искать здесь противоточный обменник было бы бесполезно.
Строение подобных сосудистых сплетений довольно хорошо изучено, однако их функция малоизвестна. Тем не менее можно высказать некоторые общие положения. У ныряющих животных очень велики как объем крови, так и ее кислородная емкость, что дает им возможность обеспечивать себя кислородом на то время, пока они находятся под водой. Во время ныряния приток крови почти ко всем частям тела прекращается; в частности, мышцы становятся практически обескровленными. Следовательно, большой объем крови животных должен в это время находиться где-то в другом месте, в каких-то вместилищах соответствующей емкости.
Такие вместилища, по всей вероятности, должны содержать артериальную, а не венозную кровь, ибо чем больше запас кислорода, тем дольше животное может находиться под водой. Наилучшим местом для этого служат не большие синусы с дряблыми стенками, а мелкие сосуды, имеющие эластичные стенки и способные поддерживать артериальное давление в течение длительных промежутков между сердечными сокращениями. Благодаря этому жизненно важные ткани — центральная нервная система и. сердечная мышца — снабжаются богатой кислородом кровью при постоянном артериальном давлении, хотя частота сердечных сокращений снижается в это время до нескольких раз в минуту.
Довольно остроумное (но в сущности маловероятное) объяснение функции артериальных сетей у китов состоит в том, что во время ныряния они помогают использовать кислород, содержащийся в окружающем их жире, поскольку растворимость кислорода {106} в жирах примерно в пять раз выше, чем в воде. Чтобы разобраться в этом, следует заняться количественной стороной этих процессов. Растворимость кислорода в физиологическом растворе при парциальном давлении, характерном для артериальной крови (примерно 100 мм рт. ст.), составляет около 0,3 мл O2 на 100 мл, а в жирах — около 1,5 мл O2 на 100 мл. Это не так уж много по сравнению с кислородной емкостью крови, равной 20—30 мл на 100 мл, то есть примерно в 20 раз выше, чем «емкость» жира. Очень небольшое количество крови вмещает в себя больше кислорода, чем большое количество жира. При этом следует помнить, что диаметр сосудов, о которых идет речь, порядка миллиметра, то есть в 100 раз больше, нем это необходимо для диффузии газов с достаточной скоростью.
А как обстоит дело с приспособлением к давлению? В строении китов есть несколько любопытных черт; некоторые из них представляют собой очевидные приспособления для плавания и ныряния, функция же других менее ясна. Весьма соблазнительно интерпретировать все и каждую необычные особенности, обнаруживаемые у китов, как приспособления, если не к плаванию и нырянию как таковым, то, во всяком случае, к высоким давлениям, которые им приходится испытывать на больших глубинах. Все признают наличие у китов некоторых анатомических структур, назначение которых нам не известно, однако, прежде чем мы сумеем выяснить их роль, мы должны попытаться объяснить, как они функционируют. Высказывая ту или иную гипотезу, нельзя ограничиваться описанием того, что нам кажется вероятным: необходимо как-то обосновать свою точку зрения.
Вернемся еще раз к обширным артериальным сетям китов, которые, как считают, помогают выдерживать давление при нырянии на большую глубину. Высказывались предположения, что они защищают головной мозг, но как это достигается? Мы знаем, что в жидкости давление равномерно распределяется во всех направлениях и что только очень прочная жесткая структура может выдержать высокое давление. Когда кит ныряет, его мягкие ткани, кровеносные сосуды, кровь и т. п. оказываются практически под {107} таким же давлением, что и окружающая вода1. Головной мозг заключен в прочный футляр — череп, который у китов очень толстый и плотный. Поэтому поддержание в черепе более низкого давления в принципе осуществимо при условии, что все имеющиеся в нем отверстия будут закрыты; если, однако, кровеносные сосуды будут открыты, то давление немедленно выровняется. Таким образом, для поддержания низкого внутричерепного давления необходимо не только прекращение кровообращения в черепе, но и наличие каких-то клапанов, которые бы не допускали поступления крови в череп, то есть обеспечивали бы поддержание постулированной разности давлений. Если при нырянии на большую глубину мозг получает кровь (а в настоящее время все согласны с этим), то внутричерепное давление должно быть равно наружному давлению.
Это вовсе не значит, что артериальные сети, расположенные вдоль позвоночника и у основания черепа, не имеют никакого отношения к нырянию и давлению; однако любая высказываемая нами гипотеза должна содержать разумное предположение не только о том, какую функцию они выполняют, но и о том, каким образом эта функция становится физически возможной.
Есть еще одна маловероятная идея, о которой мне хотелось бы упомянуть. Высказывалось мнение, что сосудистые сети в плече некоторых лазающих животных, например у ленивцев и лори, а особенно у лемуров, обусловливают способность этих животных крепко ухватываться за ветви и не выпускать их в течение длительного времени.
Авторы подобных заявлений не разъясняют, каким образом сосудистая сеть в плече обеспечивает способность мышц предплечья долгое время оставаться в состоянии сильного сокращения. Ниже этой сети сосуды вновь сливаются, образуя одну артерию; как могут мышцы знать, что кровь, которую они получают {108} по этой артерии, чем-то отличается от крови, поступающей по любой другой артерии? Единственным различием может быть несколько более низкое давление.
Возможно, венозные капилляры, располагающиеся в сети между артериальными капиллярами, остаются открытыми при повышении наружного давления во время сильного сокращения мышц, тогда как обычные вены в такой ситуации неизбежно спадаются (Саклинг, Саклинг, Уокер, 1969). А что происходит с венами, расположенными непосредственно под сосудистым пучком? Ведь если они спадутся, то сеть окажется бесполезной. Так или иначе, но сеть находится в плече, а сокращающиеся мышцы — в предплечье. Было бы полезно, если бы такого рода гипотезы давали четкое объяснение того, как действует предлагаемый механизм, и по возможности подкреплялись экспериментами, показывающими, что он в самом деле действует. Достоверным остается лишь неизбежность теплообмена в чудесной сети, а также то, что венозная кровь, возвращающаяся из конечности, погруженной в ледяную воду, пройдя через эту сеть, согревается почти до температуры артериальной крови (Шоландер и Крог, 1957). Не вызывает сомнения, что эти сети действуют как теплообменники; вопрос о том, есть ли у них какие-либо другие функции и какие именно, требует более подробного изучения.
Я уже затрагивал большинство общих черт различных противоточных механизмов. И тем не менее, рискуя показаться невыносимо многословным, вновь выделю главные пункты.
1. Противоточные умножители действуют против градиента; они требуют затраты энергии и наличия петлеобразной структуры, в которой накапливается раствор. Каналы, из которых состоит система, по размерам соответствуют капиллярам; известные примеры— образование концентрированной мочи в почках и накопление газа в плавательном пузыре.
2. Противоточные обменники действуют по нисходящему градиенту; они служат запирательными {109} приспособлениями, обеспечивающими сохранение градиентов тепла, газов и т. д. Петлеобразная структура не обязательна, но потоки должны быть направлены в противоположные стороны. Обменник может состоять из двух каналов, по которым текут разные жидкости (вода — кровь в жабрах рыб; воздух — кровь в легких птицы; кровь — кровь в плаценте), но это может быть и петля с одной жидкостью (кровь в плавнике кита, в семенниках, в мозгу).
Обменники функционируют только в тех случаях, когда возможна диффузия (тепла, кислорода, водяного пара). Диффузию по нисходящему градиенту предотвратить нельзя; эффективность обменника зависит от площади, расстояний, на которых происходит диффузия, и от других параметров (скорости течения, теплоемкости или растворимости газа и т. п.).
Для эффективного действия теплообменников кровеносные сосуды должны иметь диаметр порядка 1 мм; при такой величине сосудов газообмен минимален. Газообменники эффективны в тех случаях, когда сосуды имеют размеры капилляров (около 0,01 мм в диаметре) или при диффузии в воздухе, скорость которой в 100 000 раз выше, чем в воде.
| {110} |
В основе сравнительной физиологии лежит представление о том, что все животные более или менее сходны и поэтому их можно сравнивать между собой. Это, однако, не означает, что все они одинаковы; более того, любое отклонение от общего плана строения нередко имеет такое же важное значение и представляет не меньший интерес, чем черты сходства.
Трудность сравнения организмов разного размера иллюстрирует рис. 45, на котором изображен наш старый приятель Гулливер и житель Лилипутии, шагающий по булыжной мостовой. Когда Гулливер прибыл в Лилипутию, императору этой страны пришлось срочно решать задачу: сколько потребуется пищи, чтобы накормить Человека-Гору? Как сообщает Свифт, это составило ровно 1728 лилипутских порций. Верен ли этот расчет и на какой основе можно сравнивать млекопитающих разного размера?
Те, кому приходилось вскрывать скаковую лошадь или борзую, быть может, обратили внимание на непропорционально большие размеры сердца у этих животных. У всех млекопитающих, как правило, размеры сердца относительно общих размеров тела довольно одинаковы и вес сердца составляет примерно 5—6 г на 1 кг веса тела; мы настолько привыкли к такому соотношению, что любое отклонение от него сразу заметно.
Сходство относительных размеров сердца у большинства млекопитающих (немногим более 0,5% веса тела) на первый взгляд может показаться удивительным: ведь известно, что у мелких животных интенсивность метаболизма относительно гораздо выше, чем у крупных. Совершенно очевидно, что для достаточно интенсивного снабжения тканей кислородом {111} мало просто повысить ударный объем, который лимитируется размерами сердца. Таким образом, главная переменная, с помощью которой осуществляется необходимая регуляция, это частота сердечных сокращений, достигающая у самых мелких млекопитающих 500—1000 ударов в минуту,
Пожалуй, самое заметное различие между млекопитающими — это различие в размерах. Слон, весящий 4 тонны, в миллион раз больше землеройки,
 |
Рис. 45. Гулливер у лилипутов. Сколько ему нужно еды? |
которая весит всего 4 грамма, а самое крупное из ныне живущих млекопитающих — синий кит — больше землеройки в 25 миллионов раз. Нелегко представить 25 миллионов землероек сразу, а поскольку пища, которую они съедают за день, равна повесу примерно половине их собственной массы, то прокормить их было бы довольно трудно. Стадо из 25 слонов как-то легче себе представить, хотя это была бы тоже порядочная масса.
Если подходить к слону только с точки зрения размеров, то можно наделать серьезных ошибок. {112} Несколько лет назад в журнале Science была опубликована заметка о реакции самца слона на ЛСД. Авторы хотели изучить весьма своеобразное состояние, наблюдаемое у самцов слона и известное под названием «муст» (слово санскритского происхождения). Находящийся в состоянии муста самец становится очень свирепым и опасным, но это не имеет отношения к периоду половой активности (и вопреки распространенному мнению состояние муста не означает, что слону нужна самка). Вскоре после опубликования этой заметки в одном из писем к редактору Science расчет дозы ЛСД был назван «слоноподобной ошибкой» (Харвуд, 1963). Авторы при определении дозы ЛСД для слона исходили из количества препарата, которое приводит в ярость кошку, и, увеличив его пропорционально весу слона, сделали вывод, что ему следует ввести дозу 297 мг. Длинное описание того, чем это кончилось, можно свести к следующему: после инъекции 297 мг ЛСД (человеку этого количества хватило бы на 1500 приемов, потому что его разовая доза равна примерно 0,2 мг) слон начал трубить и метаться, а затем остановился и закачался; спустя 5 минут он упал, у него начались судороги и дефекация и он умер. Авторы пришли к выводу, что слоны отличаются особой чувствительностью к ЛСД.
Я привел этот пример для того, чтобы показать, к каким трагическим последствиям может привести недооценка проблемы масштабов. Как надо рассчитывать дозы различных лекарственных препаратов? Конечно, если мы хотим создать одинаковую концентрацию какого-либо вещества в жидкостях тела мелкого и крупного животного, то дозы должны быть пропорциональны их весу. При этом если для кошки весом 2,6 кг доза ЛСД равна 0,1 мг на 1 кг веса, то путем экстраполяции мы получаем для слона дозу 297 мг, а эта доза для него, очевидно, смертельна. Если же, как предлагает автор письма в Science, определять дозу на основе интенсивности метаболизма, то окажется, что доза для слона должна быть гораздо меньше — всего 80 мг. Это не лишено смысла, поскольку можно ожидать, что детоксикация химических веществ или их выведение из организма зависит {113} от интенсивности метаболизма. Однако следует иметь в виду различные другие соображения или особые обстоятельства. Например, возможно, что ЛСД. накапливается в головном мозгу; в этом случае положение сильно осложняется, так как при вычислении дозы пришлось бы учитывать вес мозга.
При расчете дозы ЛСД для слона можно было бы также взять за основу данные не для кошек, обладающих высокой устойчивостью к этому веществу, а, например, для человека. У человека весом 70 кг 0,2 мг ЛСД вызывают резко выраженные симптомы расстройства психики. Если рассчитать дозу для слона пропорционально весу, то она окажется равной всего 8 мг (вместо почти 300 мг). Если же исходить из интенсивности метаболизма (13 л O2 в 1 час у человека и 210 л — у слона), то доза для слона будет равна.,3 мг. Если же принять во внимание вес головного мозга (1400 г у человека и 3000 г у слона), то она снизится до 0,4 мг. Приведем для сравнения дозы ЛСД для слона (в мг), рассчитанные на основе разных критериев:
веса тела слона и эффективной дозы для кошки | 297 |
интенсивности метаболизма слона и кошки | 80 |
веса тела слона и эффективной дозы для человека | 8 |
интенсивности метаболизма слона и человека | 3 |
веса мозга слона и человека | 0,4 |
Я не намерен обсуждать здесь вопрос о том, сколько ЛСД нужно вводить слону и нужно ли вводить его вообще. Я просто хотел показать, что проблема масштабов далеко не столь проста. Однако во многих других ситуациях различия между животными гораздо более очевидны и их легче анализировать.
На рисунке 46 изображены скелеты двух животных, которых никто из нас никогда не видел живыми. Справа изображен Neohipparion — вымерший предок лошади; достаточно одного взгляда, чтобы понять, что это было животное величиной с оленя. Слева изображено другое вымершее животное — мастодонт, и хотя на рисунке он имеет такие же размеры,
| {114} |
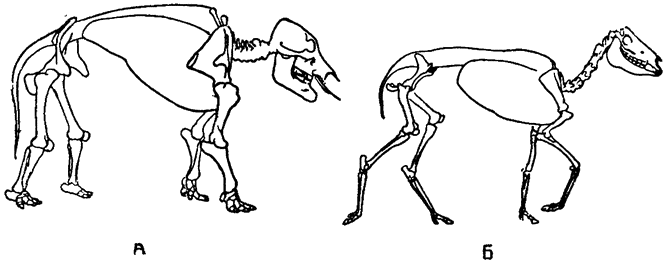 |
Рис. 46. Скелеты двух вымерших млекопитающих — Mastodon (A) и Neohipparion (Б) (по Грегори, 1912). |
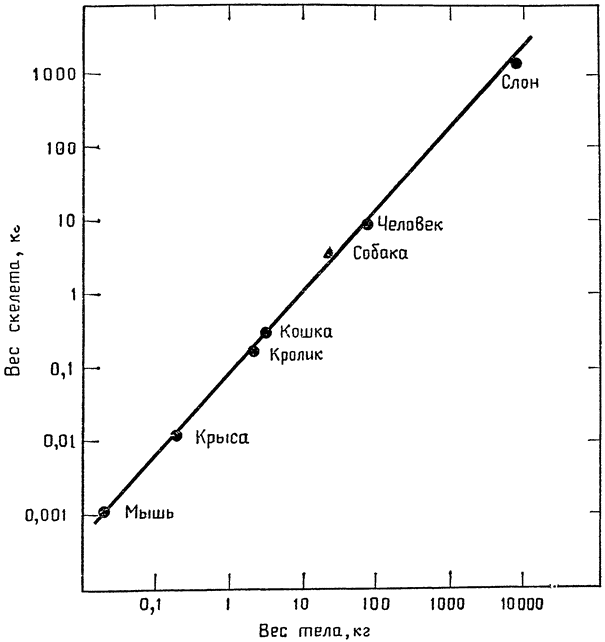 |
|
Рис. 47. Изменение веса скелета млекопитающих в зависимости от веса тела (по Кайзеру и Хейснеру, 1964). Наклон линии регрессии равен 1,13. |
| {115} |
как Neohipparion, сразу видно, что эти массивные тяжелые кости принадлежат животному величиной со слона. То обстоятельство, что мы сразу воспринимаем истинные размеры этих животных, несмотря на то, что на рисунке они имеют одинаковую величину, отражает хорошо известное правило: масса животного возрастает пропорционально кубу его линейных размеров; а для того, чтобы кости могли выдержать эту массу, площадь их поперечного сечения должна увеличиваться больше, чем она увеличивалась бы при возрастании их диаметра пропорционально линейным размерам животного. Если бы этот диаметр возрастал в линейной зависимости, то площадь сечения увеличивалась бы лишь пропорционально квадрату линейных размеров и кости сломались бы под тяжестью веса, который пропорционален кубу этих размеров.
Увеличение веса скелета с увеличением размеров у млекопитающих представлено в более общем виде на рис. 47. Этот график построен в логарифмической шкале, и все точки ложатся на одну прямую. Если бы все параметры животного изменялись в одинаковой пропорции, то вес скелета всегда составлял бы одну и ту же долю общего веса тела и соответствующий график имел бы вид прямой с наклоном 1,0. Между тем на полученном графике наклон кривой больше (1,13), то есть у крупных животных скелет относительно тяжелее.
Один из переменных физиологических параметров млекопитающих представляет для нас гораздо больший интерес, чем размеры их скелета; этот параметр — интенсивность их метаболизма. Если бы интенсивность метаболизма находилась в прямой зависимости от веса тела, то, построив график в логарифмической шкале, мы поручили бы линию регрессии с наклоном 1,0. Полная невозможность такой зависимости настолько хорошо известна, что я не стал бы еще раз упоминать об этом, если бы того не требовало дальнейшее изложение. Более 100 лет назад французские ученые высказали мнение, что у теплокровных {116} животных теплоотдача должна быть пропорциональна площади поверхности их тела и, следовательно, у мелких животных, площадь поверхности которых относительно больше, теплопродукция происходит относительно более интенсивно, чем у крупных. Аналогичные соображения лежат в основе правила Бергмана, которое гласит, что животные, обитающие в областях с холодным климатом, обычно крупнее, чем их близкие родичи, обитающие в более теплых областях (Бергман, 1847).
Впервые экспериментальную проверку этих представлений произвел Рубнер, опубликовавший соответствующие данные для собак разного размера (табл. 6).
Таблица 6
Зависимость интенсивности метаболизма от площади поверхности тела у собак (по данным Рубнера, 1883)
Вес тела, |
Площадь |
Б: А |
Интенсивность метаболизма, |
|
на 1 кг веса |
на 1 м2 |
|||
31,20 |
10 750 |
344 |
35,68 |
1036 |
24,00 |
8 805 |
366 |
40,91 |
1 112 |
19,80 |
7 500 |
379 |
45,87 |
1207 |
18,20 |
7 662 |
421 |
46,20 |
1097 |
9,61 |
5 286 |
550 |
65,16 |
1 183 |
6,50 |
3 724 |
573 |
66,07 |
1 153 |
3,19 |
2 423 |
726 |
88,07 |
1212 |
Как видно из первого столбца таблицы 6, вес собак варьировал примерно от 31 до 3 кг, то есть в диапазоне 10:1. (В настоящее время можно без труда подобрать собак, размеры которых различались бы гораздо сильнее.) Интенсивность метаболизма в расчете на 1 кг веса (четвертый столбец) по мере уменьшения веса тела повышается. Однако интенсивность метаболизма на 1 м2 поверхности (пятый столбец) изменяется очень мало, то есть отношение интенсивности метаболизма собак к площади поверхности их тела почти постоянно. Рубнер объясняет это {117} как приспособление к сохранению тепла: поскольку теплоотдача происходит с поверхности тела, она должка зависеть от площади этой поверхности. Кроме того, он весьма четко указал, что теплоотдача обусловлена не какой-то особой активностью клеток, а стимуляцией кожных рецепторов, которые в свою очередь стимулируют клетки метаболически активных тканей. Мы считаем эти доводы ошибочными.
На основании работ Рубнера был сформулирован «закон поверхности», или «правило поверхности», как я предпочитаю его называть, и площадь поверхности стали широко использовать-для оценки интенсивности метаболизма. Однако спустя всего лишь пять лет после опубликования работы Рубнера стало ясно, что теплоотдача не может быть главным фактором, обусловливающим эту зависимость. В 1888 году фон Гёсслин опубликовал результаты своих исследований, проведенных на рыбах; он установил, что у рыб потребление кислорода гораздо больше зависит от площади поверхности, чем от веса. Между тем совершенно очевидно, что рыбам нет никакой нужды сохранять тепло пропорционально площади поверхности.
После работы Гёсслина было проведено множество других исследований на эту тему, но, прежде чем заняться их рассмотрением, я уделю еще некоторое внимание поверхности тела животных. Площадь поверхности очень строго зависит от веса тела. Вместе с тем точно определить площадь поверхности тела очень трудно, и Клейбер (1967) неоднократно подчеркивает, что нельзя надеяться сделать это с точностью, превышающей 20%.
На рисунке 48 показана зависимость площади поверхности тела от веса у большого числа организмов весом от нескольких граммов до нескольких тонн. Прямая линия выражает эту зависимость для шара с удельным весом 1,0. Шар имеет наименьшую возможную поверхность по сравнению с любым другим телом равного объема, и, как видно на рисунке, площадь поверхности тела животных весьма закономерно оказывается вдвое больше, чем у шара соответствующего веса. В. правом верхнем углу рисунка имеется 7 более крупных точек, располагающихся {118} значительно выше всех остальных. Эти точки относятся не к животным, а к буковым деревьям (причины, по которым они-нанесены на график, станут ясны в дальнейшем). Относительная площадь поверхности у деревьев больше, чем у животных, но соответствующие точки ложатся на линию, параллельную линии для животных и имеющую наклон 0,67. Таким образом, в целом площадь поверхности живых организмов
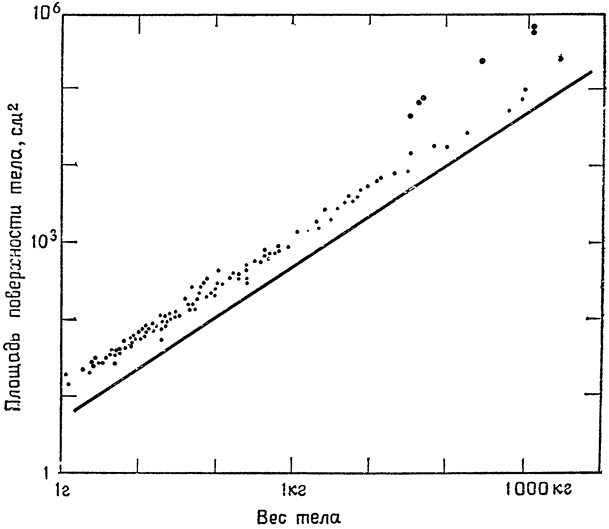 |
|
Рис. 48. Зависимость площади поверхности тела позвоночных от веса (по Хеммингсену, 1960). Сплошная линия — зависимость для шара с удельной плотностью 1,0. Крупные точки в верхнем углу — зависимость для буковых деревьев. |
удивительно строго соответствует квадрату их линейных размеров (или их объему или массе в степени 2/3). Важно понять, что сохранение строгой зависимости между площадью поверхности и объемом диктуется не одними лишь чисто геометрическими соображениями; если бы мы захотели создать функционирующую модель животного, не важно крупного или мелкого, мы бы неизбежно погрязли в необходимости все время принимать во внимание площадь {119} поверхности. В организме происходит множество процессов, зависящих от площади различных поверхностей: теплоотдача, о которой мы уже говорили, зависит от общей поверхности тела; поглощение кислорода в легких или в жабрах — от площади поверхности этих органов; диффузия кислорода через стенки капилляров — от их суммарной поверхности; всасывание питательных веществ в пищеварительном тракте — от площади всасывающей поверхности и т. д. В сущности все клетки имеют поверхности и происходящие на этих поверхностях процессы (или, как мы их теперь называем, мембранные процессы) должны зависеть от их площади.
Совершенно очевидно, что едва ли можно создать модель животного, не учитывая зависимость разных его параметров от площади поверхности тела. Приведем пример из области терморегуляции. Клейбер установил, что если бы мы создали модель быка с такой же интенсивностью метаболизма, как у мыши, с тем, чтобы она рассеивала тепло с той же скоростью, с какой оно продуцируется, то температура на поверхности модели оказалась бы гораздо выше точки кипения. И наоборот, если создать мышь, интенсивность метаболизма которой относительно веса тела соответствовала бы таковой у быка, то для того, чтобы сохранять нормальную температуру тела, ей надо было бы иметь шерстный покров толщиной не менее 20 см. Совершенно ясно, что площадь поверхности имеет важное значение и ею пренебрегать нельзя.
После опубликования работы Рубнера многие выдающиеся физиологи уделяли очень большое внимание изучению зависимости между поверхностью тела и интенсивностью метаболизма. Однако в основном ученые продолжали считать главным фактором площадь поверхности, пока Клейбер (1932) не написал статью, которая до сих пор еще оказывает большое влияние на все наши концепции в этой области. Эта статья напечатана в Hilgardia — малоизвестном сельскохозяйственном журнале, издававшемся {120} Калифорнийской экспериментальной сельскохозяйственной станцией в Дэвисе (ныне Университет штата Калифорния в Дэвисе). У меня есть редкий оттиск этой статьи с красным штампом: «Макс Клейбер. Личный экземпляр. Не вынимать из архива». Клейбер, очевидно, понял отсутствие логики в этих распоряжениях; если оттиск нельзя взять из архива, то как же его можно использовать? Вероятно, именно по этой причине он лично подарил его мне — подарок, за который я ему бесконечно благодарен.
В этой статье Клейбер показал, что интенсивность метаболизма у млекопитающих не только удивительно постоянно зависит от их веса, но и заметно отклоняется от прямой зависимости от площади поверхности. Он изучал животных самых разных размеров — от крысы до быка — и построил кривую, которую можно назвать «кривая крыса — бык». Это довольно неуклюжее название не вошло в обиход; однако вывод Клей-бера о том, что построенный в логарифмическом масштабе график зависимости интенсивности метаболизма млекопитающих от веса их тела представляет собой прямую с наклоном 0,75, заслужил всеобщее признание. Спустя два года Броди, Проктер и Эшуорс (1934) опубликовали свою хорошо известную «кривую мышь — слон», а спустя еще четыре года Бенедикт (1938) опубликовал такую же кривую в своей книге «Энергетика жизни», в которой он несколько неохотно признал, что, хотя животные ничего не знают о логарифмах, на логарифмическом графике зависимости метаболизма от веса тела точки ложатся удивительно близко к прямой линии.
Наклон прямой, построенной Броди, составил 0,734; эта величина очень близка к 0,75, постулированной Клейбером, и в сущности различие между этими двумя величинами статистически недостоверно. Рассуждения относительно значимости второго или третьего знака после запятой представляются несколько бессмысленными, если учесть, что, прежде чем Броди произвел свои расчеты, он принял некоторые «допущения» для слона. Точка, отвечающая крайним значениям размеров, оказывает на наклон линии гораздо большее влияние, чем точка, отвечающая средним размерам; в данном случае это влияние составило 30%, {121} так как «из первоначальной величины было вычтено 30% (10%, чтобы учесть стоячее положение, 20%, чтобы учесть приращение тепла за счет кормления)» (Броди, 1945).
Изучение зависимости между интенсивностью метаболизма и весом тела было проведено также на многих холоднокровных животных; полученные при этом линии регрессии имели такой же наклон, хотя располагались ниже. Сходный наклон имеет линия регрессии даже для буковых деревьев. Это заставляет навсегда отказаться от представления о терморегуляции как о главном факторе, определяющем постоянство соотношения между продукцией тепла и весом тела. Для многих беспозвоночных зависимость между интенсивностью метаболизма и весом тела также описывается прямыми линиями, которые статистически имеют такой же наклон. У некоторых животных, однако, наклон линий регрессии сильно отличается, а у ряда моллюсков и насекомых он равен 1,0, то есть интенсивность их метаболизма прямо пропорциональна весу тела.
Подробный обзор всех этих данных произвел Хеммингсен (1960), рассмотревший очень разных живых существ — от микроорганизмов до самых крупных млекопитающих (рис. 49). О широте такого диапазона свидетельствует уже одно то, что каждое деление на координатных осях соответствует разнице не в 10, а в 1000 раз. На графике точки для теплокровных хорошо ложатся на прямую; сходная прямая, но лежащая несколько ниже, получается для холоднокровных животных, а на продолжение ее нижнего конца укладываются точки для многих беспозвоночных. Данные по микроорганизмам тоже образуют прямую с таким же наклоном, как и две предыдущие прямые. Однако наклоны всех этих трех прямых заметно отличаются от линии, описывающей зависимость от площади поверхности (прерывистая линия с наклоном 0,67), и от линии, описывающей прямую зависимость от веса (прерывистая линия с наклоном 1,00).
Все три интересующие нас линии описываются хорошо известным экспоненциальным уравнением
у = b · ха
| {122} |
В логарифмической форме это уравнение превращается в линейное:
log y = а · log х + log b,
где а — наклон прямой — соответствует показателю степени в предыдущем уравнении. Аналогично если мы обратимся к кривым, описывающим процесс метаболизма, которые мы уже обсуждали, то мы получим,
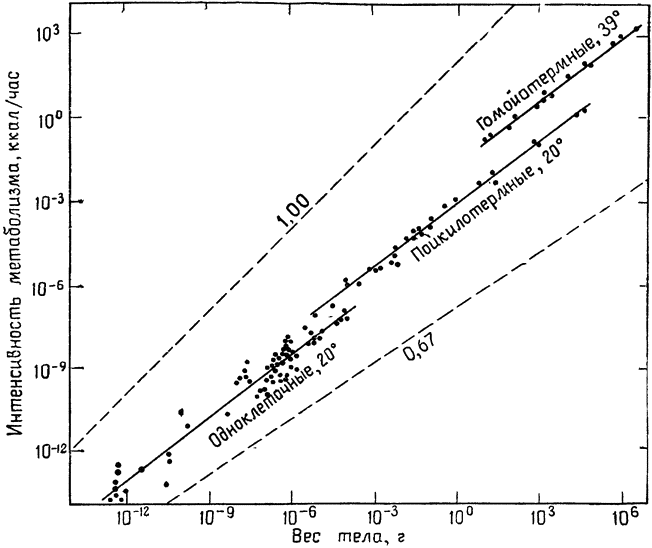 |
Рис. 49. Зависимость интенсивности метаболизма от веса у разных организмов (по Хеммингсену, 1960). |
что логарифм интенсивности метаболизма связан с логарифмом веса тела линейной зависимостью
log (интенсивность метаболизма) = а · log (вес тела) + k.
Вернемся теперь к обсуждению биологического смысла коэффициентов а, то есть наклонов прямых, представляющих собой линии регрессии, и статистической достоверности различий в их значениях. Отличается ли наклон 0,75, на котором настаивает Клейбер, достоверным образом от величины 0,67? Ответ состоит в том, что для установления достоверности {123} этого различия надо рассматривать животных, размеры которых отличаются больше чем в 9 раз. Напомним, что диапазон различий в размерах между собаками, с которыми работал Рубнер, был примерно таким. Поэтому Рубнер, в сущности, не располагал достаточными данными, которые позволили бы ему достоверным образом установить отличие величины наклона от значения 0,67, связанного просто с отношением поверхности к объему. В тех работах, которые проводились на большом числе млекопитающих очень разных размеров, величина а всегда оказывалась больше, чем 0,67. Например, Броди и Проктер (1932) получили значение 0,734, а Клейбер (1961) — 0,756. Клейбер показал, что эти величины можно считать достоверно отличными от 0,75 лишь в том случае, если вес изучаемых животных будет варьировать от менее чем 4 г до более чем 800 т. Однако известно, что самое крупное из ныне живущих животных — синий кит — весит лишь около 100 т, и поэтому я полагаю, что мы никогда не будем располагать фактическими данными, которые бы позволили достоверно установить отличие величины а от 0,75. Один из доводов, приводимых Клейбером в пользу значения 0,75 (3/4), состоит в том, что с этой величиной гораздо проще оперировать при вычислениях.
Повторим вкратце те этапы, которые привели к нашим современным представлениям в этой области, В начале XIX века французские ученые пришли к выводу, что теплоотдача должна зависеть от площади поверхности животного. Рубнер впервые изучил этот вопрос экспериментально на собаках, очень сильно различающихся по размеру. Он обнаружил весьма тесную зависимость интенсивности метаболизма от площади поверхности тела, после чего соотношение этих величин стали использовать в качестве основного эталона во всех исследованиях метаболизма. Клейбер на основании изучения данных об интенсивности метаболизма у млекопитающих, различающихся по размерам гораздо сильнее, пришел к выводу, что соответствующий показатель весьма близок к весу тела в степени 3/4.
В вводной статье к «Ежегодному обзору физиологии» (Annual Review of Physiology) за 1967 год Клейбер {124} не без юмора подчеркивает универсальность этой величины. В этой статье, озаглавленной «Старый профессор животноводства пережевывает», он коснулся ряда интересных проблем, и в том числе физиологии жителей Лилипутии. Сделав некоторые допущения о весе лилипутов, Клейбер установил, что при определении количества пищи для Гулливера они приняли наклон линии регрессии равным 0,76; эта величина, по существу, не отличается от той, которую указывал Клейбер в своей статье, опубликованной в журнале Hilgardia в 1932 году, и тем самым лилипуты опередили его на 233 года. Клейбер указывает также, что если бы лилипуты основывали свои расчеты на «правиле поверхности» Рубнера, то Гулливер получил бы всего 675 порций и ему пришлось бы голодать1.
Можем ли мы проанализировать, что в сущности означает этот столь часто встречающийся наклон 0,75? Прежде чем попытаться ответить на этот вопрос, рассмотрим некоторые трудности, возникающие при расчетах, касающихся живых организмов. Рис. 50 дает наглядное представление о том, сколь быстро возрастает интенсивность метаболизма по мере уменьшения веса тела у млекопитающих. Резкое возрастание метаболизма у самых мелких животных означает, что снабжение кислородом и питательными веществами у них должно происходить в 100 раз более интенсивно, чем у крупных животных. Рассмотрим сначала факторы, от которых зависит снабжение тканей кислородом, а затем займемся газообменом в легких, ибо и то и другое играет важную роль в снабжении организма кислородом. {125}
Плотность распределения капилляров (число капилляров, приходящихся на единицу площади поперечного сечения ткани). Скорость диффузии кислорода из капилляра в метаболически активную клетку определяется: 1) расстоянием, на котором происходит диффузия; 2) разностью между парциальным давлением кислорода в капилляре и в клетке. Первое условие зависит просто от плотности распределения
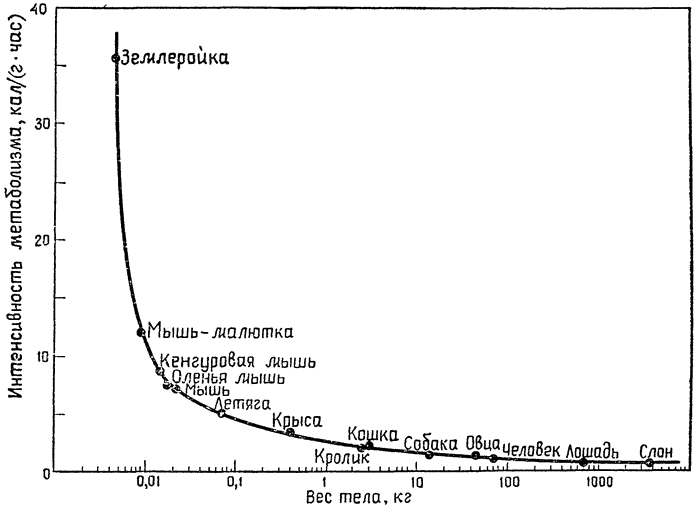 |
Рис. 50. Зависимость интенсивности метаболизма (в пересчете на 1 г веса тела) от общего веса для разных млекопитающих. |
капилляров, тогда как на второе влияет кривая диссоциации кислорода в крови, а также эффект Бора; для того чтобы эффект Бора мог осуществиться за то короткое время, пока кровь остается в капиллярах, необходимо присутствие карбоангидразы.
Крог (1929) понимал необходимость высокой плотности распределения капилляров в тканях мелких животных, и он подтвердил это, произведя подсчет числа капилляров, приходящихся на единицу площади поперечного сечения мышцы у лошади, овцы и морской свинки. Примерно такие же данные получили впоследствии некоторые другие ученые, однако различия {126} в использованных ими методах не дают возможности провести надежные сравнения. Поэтому мы подсчитали число капилляров в разных мышцах у ряда млекопитающих, применяя во всех случаях один и тот же метод. Оказалось, что число капилляров в разных мышцах одного и того же животного значительно варьирует, причем эти вариации лишь частично удается связать с соотношением красных и белых волокон в мышцах; оказалось также, что плотность
 |
Рис. 51. Плотность распределения капилляров в икроножной мышце у млекопитающих разного размера (по Шмидту-Нильсену и Пенникуику, 1961). |
распределения капилляров гораздо менее тесно связана с общими размерами животного, чем предполагалось (рис. 51), Правда, у самых мелких животных плотность распределения капилляров действительно была очень высокой, однако у более крупных видов, начиная от кролика и выше, определённой зависимости числа капилляров от веса обнаружить не удалось. Насколько я могу судить, это должно означать, что какие-то другие соотношения играют более важную роль, чем расстояние, на котором происходит диффузия. В мышце капилляры находятся среди мышечных {127} волокон, которые в любом случае представляют собой главную функциональную единицу мышцы. Быть может, определять относительные размеры мышечного волокна у крупного животного невозможно по причинам, связанным с величиной сократительных элементов или с условиями проведения потенциала действия. В таком случае изменения плотности распределения капилляров в мышцах крупных млекопитающих окажутся ограниченными.
Давление полунасыщения. Рассмотрим теперь давление полунасыщения кислорода в наполняющей капилляры крови, описываемое кривой диссоциации оксигемоглобина (рис. 52). Здесь нас интересует цельная, интактная кровь при нормальном для данного организма рН, потому что по кровеносным сосудам течет именно цельная кровь, а не разбавленные растворы гемоглобина в фосфатном буфере. Общая тенденция не вызывает сомнений: кривые диссоциации крупных животных располагаются левее, мелких — правее. Давление полунасыщения (P50), иногда называют «напряжением отдачи» (unloading tension), однако нам не нужно точно определять величину этого давления, чтобы убедиться, что у мелких животных гемоглобин отдает кислород при более низком парциальном давлении, чем у крупных. На рис. 52 показано также влияние кислоты на кривую диссоциации оксигемоглобина у мыши; наблюдаемый сдвиг кривой вправо (эффект Бора) повышает давление полунасыщения.
После того как была установлена зависимость между кривой диссоциации оксигемоглобина и общими размерами тела у млекопитающих, которая была истолкована как отражение большей потребности мелких животных в кислороде, интерес к исследованиям на цельной крови сильно возрос и было описано много других аналогичных примеров среди млекопитающих. Особый интерес ввиду своих больших размеров представляет слон, кривая диссоциации которого располагается левее кривых для всех других млекопитающих.
Эффект Бора. Как уже было сказано выше, эффект Бора весьма полезен, поскольку он повышает давление полунасыщения кислорода. На рис. 53 собраны
| {128} |
 |
|
Рис. 52. Кривые диссоциации оксигемоглобина для ряда млекопитающих (по Шмидту-Нильсену и Ларимеру, 1958). 1 — слон (по Бартельсу и др., 1963); 2 — лошадь; 3 — человек; 4 — овца; 5 — лисица; 6 — кошка; 7 — крыса; 8 — мышь. Прерывистая кривая (8') показывает действие кислоты (эффект Бора) на кровь мыши (кривая 8). |
 |
Рис. 53. Действие кислоты на отдачу кислорода из крови (эффект Бора, выражаемый как Δlog P50/ΔрН) проявляется сильнее у мелких, чем у крупных животных (по Риггсу, 1960). |
| {129} |
данные об эффекте Бора у разных млекопитающих. Как показывает этот рисунок, эффект Бора сильно возрастает но мере увеличения веса животного — от слона к мыши; подкисление крови больше повышает утилизацию кислорода у мелких животных. Среди всех приведенных на рисунке животных обращает на себя внимание чихуахуа, точка для которой сильно отклоняется в сторону и не укладывается на линию регрессии. Быть может, то обстоятельство, что эффект Бора для этого животного не. отличается от такового для обычной собаки, свидетельствует о том, что, несмотря на небольшой вес и своеобразный внешний вид, чихуахуа все же остается собакой.
Карбоангидраза. Для того чтобы эффект Бора усилил отдачу кислорода тканям, совершенно необходимо, чтобы кровь была подкислена до того, как она выйдет из капилляра. Эритроцит находится в капилляре лишь в течение какой-то доли секунды; за такое короткое время СO2 тканей не успела бы превратиться в угольную кислоту, если бы не присутствие карбо-ангидразы, ускоряющей реакцию гидратации СO2. Мы с Ларимером исследовали содержание карбо-ангидразы в эритроцитах разных млекопитающих и обнаружили, что у мелких животных содержание этого фермента в эритроцитах гораздо выше, чем у крупных. Считается, что карбоангидраза не играет существенной роли в процессе дыхания; блокирование ее функции при помощи диамокса не оказывает заметного влияния. Давенпорт описал карбоангидразу как фермент, не имеющий определенной функции. Однако мы считаем, что карбоангидраза, не будучи необходимой для транспорта СО2, играет важную роль, способствуя адекватной отдаче кислорода тканям; подобная функция карбоангидразы, очевидно, должна иметь особенно большое значение во время физической нагрузки (Лаример и Шмидт-Нильсен, 1960),
А теперь рассмотрим вкратце противоположный элемент системы переноса кислорода — поглощение кислорода в легких, В этом случае вычисление {130} соотношений гораздо проще и мы можем провести более глубокий анализ.
Прежде всего оказывается, что у млекопитающих объем легких находится в прямой зависимости от веса тела (рис. 54). Если нанести на ось абсцисс логарифмы веса тела различных млекопитающих, а
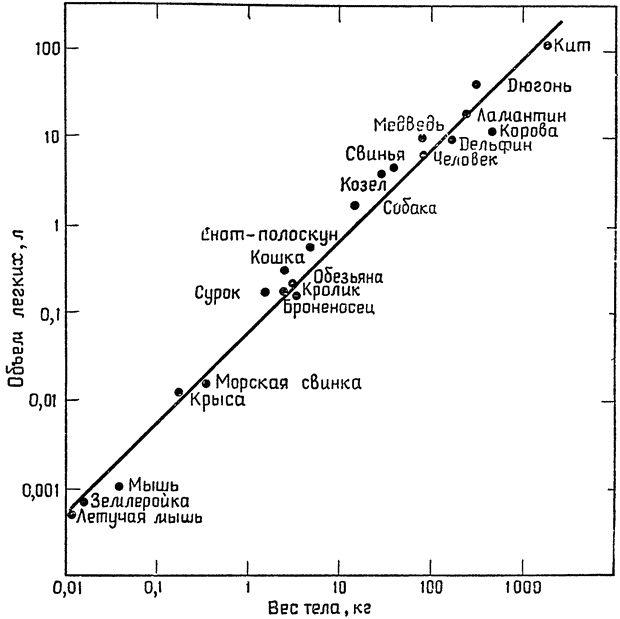 |
|
Рис. 54. Зависимость объема легких у млекопитающих от веса тела (по Теннею и Реммерсу, 1963). Наклон линии регрессии равен 1,02. |
на ось ординат — логарифмы объема их легких, то получится прямая линия с наклоном, очень близким к 1; иными словами, относительный объем легких у всех млекопитающих одинаков и составляет примерно 6,3% их общего объема. Как уже отмечалось, всегда имеются некоторые отклонения от таких линий регрессии. Эти отклонения могут быть обусловлены ошибками измерения или различиями в методах исследования, однако нельзя забывать, что иногда {131} отклонения представляют особый интерес и заслуживают большего внимания, чем типичная картина, после того как она достаточно хорошо выяснена.
Если обратиться к диффузионной поверхности легкого, то оказывается, что эта переменная непосредственно зависит от скорости поглощения кислорода, а не от размеров тела животного (рис. 55). С физиологической
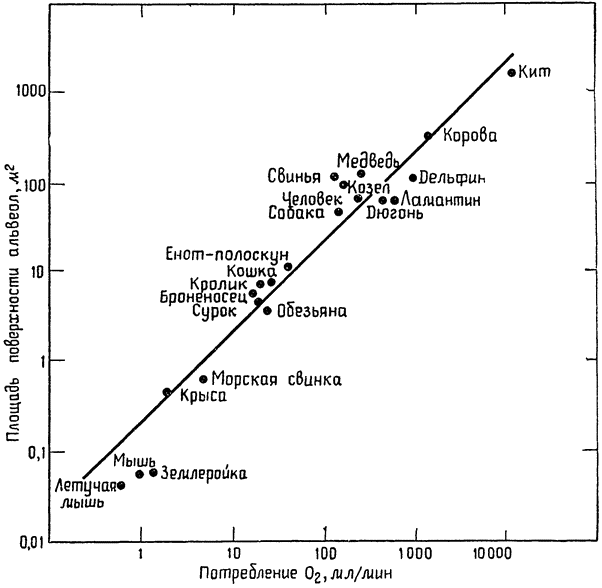 |
Рис. 55. Зависимость площади диффузионной поверхности легких у млекопитающих от скорости потребления кислорода (по Теннею и Реммерсу, 1963). |
точки зрения это понятно. Поглощение кислорода, конечно, пропорционально весу тела в степени 3/4, и поэтому площадь диффузионной поверхности связана с ним такой же зависимостью. Таким образом, принимая во внимание соотношение с размерами тела, можно сделать вывод, что у млекопитающих объем легких и их диффузионная поверхность действительно соответствуют потребностям организма в кислороде. {132}
Как показывает следующий пример, применимость аллометрических уравнений и их предсказательная сила могут оказаться гораздо шире. Жизненная емкость легких1 млекопитающих находится примерно в такой же зависимости от веса тела, как объем легких (см. рис. 54), и описывается уравнением
Жизненная емкость = 0,053 В1,0,
где В — вес тела.
Объем дыхательного воздуха также зависит от веса тела:
Дыхательный воздух = 0,0063 В1,0.
Если разделить дыхательный воздух на жизненную емкость, то мы получим безразмерную величину 0,1. Таким образом, можно сделать вывод, что у млекопитающих объем дыхательного воздуха соответствует 0,1 жизненной емкости легких независимо от веса тела. Хотя от этого общего правила бывают отклонения, оно полезно, так как дает возможность предсказывать физиологические параметры тех или иных животных, например крысы или лошади; кроме того, оно служит неким эталоном, с которым можно сопоставлять результаты конкретных исследований. В данном случае мы получили в итоге безразмерную величину, которая свидетельствует о сходстве вентиляции легких у всех млекопитающих. В этом отношении все млекопитающие подобны друг другу без каких-либо специфических зависимостей от размеров тела.
Несколько лет назад Дрорбаф (1960), подробно изучая физиологию мышей, крыс, кроликов и собак, в числе других параметров рассматривал податливость легких. Он установил, что она непосредственно зависит от веса тела и описывается уравнением
Податливость = 0,00121 B1,0 мл (см H2O)–1.
Как показывает это уравнение, число кубических миллиметров, на которые растягивается легкое при изменении давления воды на 1 см, прямо зависит от {133} веса тела животного, и поэтому податливость легких крупного животного определяют просто относительно его размеров. Таким же образом зависит от размеров тела и жизненная емкость легких. Разделив одно уравнение на другое, мы получим удельную податливость:
Удельная податливость = 0,019 (см Н2O).
В результате этих преобразований мы приходим к выражению, в котором вес тела отсутствует; таким образом, удельная податливость не зависит от размеров животного. Полученное соотношение означает, что изменение давления на 1 см Н2O вызывает у всех млекопитающих одинаковое изменение объема легких относительно их жизненной емкости; это изменение составляет 1,9% объема. На основании этих соображений Дрорбаф высказал верное предположение, что давление при вдохе каждой порции воздуха должно быть одинаковым у всех млекопитающих.
Определение величин множества физиологических переменных и выведение безразмерных величин, выражающих соотношения между этими переменными, приобретают все более важное значение. В свое время Адольф (1949) опубликовал интересную работу по этим вопросам, а несколько лет назад Уолтер Сталь, незадолго до своей смерти, завершил работу над большой монографией «Физиологическое сходство и моделирование».
Прежде чем закончить эту главу, мне кажется уместным еще раз напомнить, что впервые применение экспоненциальных уравнений для исследования зависимости физиологических функций от размеров тела ввел Клейбер, который в 1932 году произвел четкий анализ интенсивности метаболизма. Это избавило нас от необходимости связывать интенсивность метаболизма с площадью поверхности тела и прекратило горячие дебаты о том, как определять «свободную» или «истинную» поверхность. «Закон поверхности» как таковой не сохранился даже в виде «правила поверхности», а анализ различных функций относительно размеров тела стал одним из интереснейших подходов к изучению того, как работает организм животных.
| {134} |
Adolph Е. F., Quantitative relations in the physiological constitutions of mammals, Science, 109, 579—585, 1949.
Albers C, Der Mechanismus des Warmehechelns beim Hund. I. Die Ventilation und die arteriellen Blutgase wahrend des Warmehechelns, Pflugers Arch, ges Physiol, 274, 125—147, 1961.
Baker M. A., Hayward J. N., The influence of the nasal mucosa and the carotid rete upon hypothalamic temperature in sheep, J. Physiol., 198, 561—579, 1968.
Banko W. E., The Trumpeter Swan. North American Fauna, № 63, 1960.
Barnett С H., Harrison R. J., Tomlinson J. D. W., Variations in the venous system of mammals, Biol. Rev., 33, 442—487, 1958.
Bartels H., Hilpert P., Barbey K., Betke K., Riegel K., Lang E. M, Metcalfe J., Respiratory functions of blood of the yak, llama, camel, Dybowski deer, and African elephant, Am. J. Physiol., 205, 331—336, 1963.
Benedict F. G., Vital Energetics. A Study in Comparative Basal Metabolism, 1938.
Berg Т., Steen J. В., The mechanism of oxygen concentration in the swim-bladder of the eel, J. Physiol., 195, 631—638, 1968.
Bergmann C, Ueber die Verhaltnisse der Warmeokonomie der Thiere zu ihrer Grosse, Gottinger Studien, 595—708, 1847.
Brett J. R., The relation of size to rate of oxygen consumption and sustained swimming speed of sockeye salmon (Oncorhynchus nerka), J. Fish. Res. Bd. Can., 22, 1491—1501, 1965.
Bretz W. L., Schmidt-Nielsen, K., Patterns of air flow in the duck lung, Fedn. Proc, 29, 662, 1970.
Bretz W. L., Schmidt-Nielsen K., Bird respiration: flow patterns in the duck lung, J. exp. Biol., 54, 103—118, 1971.
Brody S., Bioenergetics and Growth. With Special Reference to the Efficiency Complex in Domestic Animals, New York, 1945.
Brody S., Procter R. C, Relation between basal metabolism and mature body weight in different species of mammals and birds, Missouri Agr. Exp. Sta. Res. Bull., 166, 89—101, 1932.
Brody S., Procter R. C., Ashworth U. S., Basal metabolism, endogenous nitrogen, creatinine and neutral-sulphur excretions as functions of body weight, Missouri Arg. Exp. Sta. Res. Bull., 220, 1—40, 1934.
Calder W. A., Schmidt-Nielsen K., Evaporative cooling and respiratory alkalosis in the pigeon, Proc. natn. Acad. Sci. U. S. A., 55, 750—756, 1966. {135}
Calder W. A., Schmidt-Nielsen K., Panting and blood carbon dioxide in birds, Am. J. Physiol., 215, 506—508, 1968.
Carey F. G., Teal J. M., Heat conservation in tuna fish muscle, Proc. natn. Acad. Sci. U. S. A., 56, 1464—1469, 1966.
Carey F. G., Teal J. M., Mako and porbeagle: warm-bodied sharks, Сотр. Biochem. Physiol, 28, 199—204, 1969.
Collins J. C, Pilkington T. C, Schmidt-Nielsen K., A model of respiratory heat transfer in a small mammal, Biophys. J., 11, 886—914, 1971.
Crawford E. C, Mechanical aspects of panting in dogs, J. appl. Physiol., 17, 249—251, 1962.
Crawford E. C, Kampe G., Resonant panting in pigeons, Сотр. Biochem. Physiol., 40A, 549—552, 1971.
Daniel P. M., Dawes J. D. K., Pnchard M. M. L., Studies on the carotid rete and its associated arteries, Phil. Trans. Roy. Soc, B, 237, 173—208, 1953.
Drorbaugh J. E., Pulmonary function in different animals, J. appl. Physiol., 15, 1069—1072, 1960.
Fedde M. R., Burger R. E., Kitchell R. L., Electromyographic studies on certain respiratory muscles of the chicken, Poult. Sci., 42, 1269, 1963.
Gifford W. E., McMahon H. O., A new low-temperature gas expansion cycle, Part II, Adv. in Cryog. Eng., 5, 368—372, 1960.
Gjonnes В., Schmidt-Nielsen K., Respiratory characteristics of kangaroo rat blood, J. cell. сотр. Physiol., 39, 147—152, 1952.
Gregory W. K., Notes on the principles of quadrupedal locomotion and on the mechanism of the limbs in hoofed animals, Ann. N. Y. Acad. Sci., 22, 267—294, 1912.
Grigg G. C, Water flow through the gills of Port Jackson sharks, J. ex p. Biol., 52, 565—568, 1970.
Haies J. R. S., The partition of respiratory ventilation of the panting ox, J. Physiol., 188, 45—46P, 1966.
Hales J. R. S., Bligh J., Respiratory responses of the conscious dog to severe heat stress, Experientia, 25, 818—819, 1969.
Hales J. R. S., Findlay J. D., Respiration of the ox: normal values and the effects of exposure to hot environments, Resp. Physiol, 4, 333—352, 1968.
Hall F. G., Dill D. В., Guzman Barron E. S., Comparative physiology in high altitudes, J. cell сотр. Physiol, 8, 301—313, 1936.
Harwood P. D., Therapeutic dosage in small and large mammals. Letter to the Editor, Science, 139, 684—685, 1963.
Hemmingsen A. M., Energy metabolism as related to body size and respiratory surfaces, and its evolution, Reports of the Steno Memorial Hospital and Nordisk Insulinlaboratorium, 9, 1 — 110, 1960.
Hinds D. S., Calder W. A., Tracheal dead space in the respiration of birds, Evolution, 25, 429—440, 1971.
Hutchens J. O., Podolsky В., Morales M. F., Studies on the kinetics and energetics of carbon and nitrogen metabolism of Chilomonas Paramecium, J. cell comp. Physiol., 32, 117—141, 1948. Jackson D. C, Schmidt-Nielsen K., Countercurrent heat exchange in the respiratory passages, Proc. natn. Acad. Sci. U. S. A., 51, 1192—1197, 1964. {136}
Jenkin F., Gas- and Caloric-Engines; в кн. «Heat and its Mechanical Applications», London, 101—174, 1885.
Kayser Ch., Heusner A., Etude comparative du metabolismc energetique dans la serie animale, J. Physiol, 56, 489—524, 1964.
King A. S., Structural and functional aspects of the avian lungs and air sacs, Int. Rev. Gen Exp. Zool., 2, 171—267, 1966.
King A. S., Cowie A. F., The functional anatomy of the bronchial muscle of the bird, J. Anat., 105, 323—336, 1969.
Kleiber M., Body size and metabolism, Hilgatdia, 6, 315—353, 1932.
Kleiber M., The Fire of Life. An Introduction to Animal Energetics, New York, 1961.
Kleiber M., Prefatory chapter: An old professor of animal husbandry ruminates, Ann. Rev. Physiol, 29, 1—20, 1967.
Krogh A., The Anatomy and Physiology of Capillaries, New Haven, 1929.
Kuhn W., Ramel A., Kuhn H. J., Marti E., The filling mechanism of the swimbladder. Generation of high gas pressures through hairpin countercurrent multiplication, Experientia, 19, 497—511, 1963.
Landes R. R., Leonhardt К. О., Duruman N., A clinical study of the oxygen tension of the urine and renal structures. II, J. Urol, 92, 171—178, 1964.
Larimer J. L., Schmidt-Nielsen K., A comparison of blood carbonic anhydrase of various mammals, Сотр. Biochem. Physiol, 1, 19—23, 1960.
Lasiewski R. C, Calder W. A., A preliminary allometric analysis of respiratory variables in resting birds, Resp. Physiol, 11, 152—166, 1971.
Lasiewski R. C, Dawson W. R., A re-examination of the relation between standard metabolic rate and body weight in birds, Condor, 69, 13—23, 1967.
Lasiewski R. C, Snyder G. K., Responses to high temperature in nestling double-crested and pelagic cormorants, Auk, 86, 529—540, 1969.
Матюхин В. А., Столбов А. Я., Потребление кислорода байкальским омулем (Coregonus autumnalis migratorius) и хариусом (Thymallus arcticus P.) в зависимости от скорости плавания; в кн. «Адаптация к высокогорью и гипоксия. Адаптация водных животных», Новосибирск, АН СССР, Сибирское отделение, 147—149, 1970.
Матюхин В. А., Энергетический обмен и к. п. д. пресноводных рыб при различных температурах и дозированных двигательных нагрузках; в кн. «Адаптация к высокогорью и гипоксия. Адаптация водных животных», Новосибирск, АН СССР, Сибирское отделение, 149—152, 1970.
Murrish D. Е., Schmidt-Nielsen К., Exhaled air temperature and water conrservation in lizards, Resp. Physiol, 10, 151—158, 1970.
Naya J. K., Van Handel E., The fuel for sustained mosquito flight, J. Insect Physiol, 17, 471—481, 1971.
Nielsen В., Nielsen M., Body temperature during work at different environmental temperatures, Acta physiol scand., 56, 120—129, 1962. {137}
Norris K. S., The ecology of the desert iguana, Dipsosaurus dorsa-lis, Ecology, 34, 265—287, 1953.
Odum E. P., Rogers D. Т., Hicks D. L., Homeostasis of the nonfat components of migrating birds, Science, 143, 1037—1039, 1964.
Prange H. D., Schmidt-Nielsen K., The metabolic cost of swimming in ducks, J. exp. Biol., 53, 763—777, 1970.
Raab J., Schmidt-Nielsen K., Effect of activity on water balance of rodents, Fedn. Proc, 30, 371, 1971.
Rayner M. D., Keenan M. J., Role of red and white muscles in the swimming of the skipjack tuna, Nature, 214, 392—393, 1967.
Riggs A., The nature and significance of the Borh effect in mammalian hemoglobins, J. gen. Physiol, 43, 737—752, 1960.
Rikmenspoel R., Sinton S., Janick J. J., Energy conversion in bull sperm flagella, J. gen. Physiol, 54, 782—805, 1969.
Rothschild Lord, The movements of spermatozoa; в кн. «Mammalian Germ Cells», Siba Foundation Symposium, London, 1953, 122—133.
Rubner E., Ueber den Einfluss der Korpergrosse auf Stoff-und Kraftwechsel. Z. Biol, 19, 535—562, 1883.
Salt G. W., Respiratory evaporation in birds, Biol. Rev., 39, 113—136, 1964.
Scheid P., Piiper J., Analysis of gas exchange in the avian lung: theory and experiments in the domestic fowl, Resp. Physiol, 9, 246—262, 1970.
Шмидт-Нильсен К., Животные пустынь, Физиологические проблемы тепла и воды, изд-во «Наука», Л., 1972.
Schmidt-Nielsen К., Animal Physiology, Englewood Cliffs, 1970.
Schmidt-Nielsen К., Bretz W. L., Taylor C. R., Panting in dogs: unidirectional air flow over evaporative surfaces, Science, 169, 1102—1104, 1970, a.
Schmidt-Nielsen K., Hainsworth F. R., Murrish D. E., Countercurrent heat exchange in the respiratory passage: effect on watt and heat balance, Resp. Physiol, 9, 263—276, 1970, 6.
Schmidt-Nielsen K., Kanwisher J., Lasiewski R. C, Cohn J. E., Bretz W. L., Temperature regulation and respiration in the ostrich, Condor, 71, 341—352, 1969.
Schmidt-Nielsen K., Larimer J. L., Oxygen dissociation curves of mammalian blood in relation to body size, Am. J. Physiol, 195, 424—428, 1958.
Schmidt-Nielsen K., Pennycuik P., Capillary density in mammals in relation to body size and oxygen consumption, Am. J. Physiol, 200, 746—750, 1961.
Schmidt-Nielsen K., Schmidt-Nielsen В., Jarnum S. A., Houpt T. R., Body temperature of the camel and its relation to water economy, Am. J. Physiol, 188, 103—112, 1957.
Scholander P. F., Counter current exchange. A principle in biology, Hvalradets Skrifter, № 44, 1—24, 1958.
Scholander P. F., Van Dam L., Secretion of gases against high pressures in the swimbladder of deep-sea fishes. I. Oxygen dissociation in blood, Biol. Bull, 107, 247—259, 1954.
Scholander P. F., Krog J., Countercurrent heat exchange and vascular bundles in sloths, J. appl. Physiol, 10, 405—411, 1957. {138}
Soum J. M., Recherches physiologiques sur lappareil respiratoire des oiseaux, Ann. Univ. Lyon, 28, 1—126, 1896.
Stahl W. R., Physiological Similarity and Modeling. The Application of Dimensional Analysis and Physical Similarity Theory to Mammalian Physiology, New Yok, 1972.
Steno N., De musculi et glandulis. Amstelodami. Quoted from Broman, I (1921), Z. ges. Anat., 60, 439, 1664.
Suckling J. A., Suckling E. E., Walker A., Suggested function of the vascular bundles in the limbs of Perodicticus potto, Nature, 221, 379—380, 1969.
Свифт Дж., Путешествия в некоторые отдаленные страны света Лемюэля Гулливера, сначала хирурга, а потом капитана нескольких кораблей, М., изд-во «Художественная литература», 1967.
Taylor С. R., The eland and the oryx, Sci. Am., 220, 88—95, 1969.
Taylor C. R., Dehydration and heat: effects on temperature regulation of East African ungulates, Am. J. Physiol., 219, 1136—1139, 1970.
Taylor С. R., Dmiel R., Fedar M., Schmidt-Nielsen K., Energetic cost of running and heat balance in a large bird, the rhea, Am. J. Physiol., 221, 597—601, 1971, a.
Taylor С. R., Schmidt-Nielsen K., Dmiel R., Fedak M., Effect of hyperthermia on heat balance during running in the African Hunting Dog, Am. J. Physiol., 220, 823—827, 1971, 6.
Taylor C. R., Schmidt-Nielsen K., Raab J. L., Scaling of the energetic cost of running to body size in mammals, Am. J. Physiol., 219, 1104—1107, 1970.
Tennent D. M., A study of water losses through the skin in the rat, Am. J. Physiol., 145, 436—440, 1946.
Tenney S. M., Remmers J. E., Comparative quantitative morphology of the mammalian lung: diffusing area, Nature, 197, 54—56, 1963.
Thomas S. P., Suthers R. A., Oxygen consumption and physiological responses during flight in an echolocating bat, Fedn. Proc, 29, 265, 1970.
Tucker V. A., Respiratory physiology of house sparrows in relation to high-altitude flight, J. exp. Biol., 48, 55—56, 1968, a.
Tucker V. A., Respiratory exchange and evaporative water loss in the flying budgerigar, J. exp. Biol., 48, 67—87, 1968,6.
Tucker V. A., Energetic cost of locomotion in animals,, Сотр. Bio-chem. Physiol, 34, 841—846, 1970.
Tucker V. A., Flight energetics in birds, Am. Zool., 11, 115—124, 1971.
Verzar F., Keith J., Parchet V., Temperatur und Feuchtigkeit der Luft in den Atemwegen, Pflugers Arch. ges. Physiol, 257, 400—416, 1953.
Von Hoesslin H., Ueber die Ursache der scheinbaren Abhangigkeit des Umsatzes von der Gresse der Korperoberflache. Du Bois Reymond Arch. Anat. Physiol, 323—379, 1888.
Walker J. E. C, Wells R. E., -Merrill E. W., Heat and water exchange in the respiratory tract, Am. J. Med., 30, 259—267, 1961. {139}
Webb P., Air temperatures in respiratory tracts of resting subjects in cold, J. appl. Physiol., 4, 378—382, 1951. Webb P., Respiratory heat loss in cold, Fedn. Proc, 14, 486, 1955.
Weis-Fogh Т., Weight economy of flying insects, Trans. Ninth Int. Congr. Ent., 1, 341—347, 1952.
Wohlschlag D. E., Cameron J. N., Cech J. J., Seasonal changes in the respiratory metabolism of the pinfish (Lagodon rhomboides), Contr. Marine Sci., 13, 89—104, 1968. Zeuthen E., The ventilation of the respiratory tract in birds, Kl. danske Vidensk. Selsk. Biol. Medd, 17, 1—50, 1942.
| {140} |
1 Но это, конечно, не вечный двигатель, так как система работает лишь в том случае, когда нижний конец большого цилиндра постоянно нагревается за счет поступающего извне тепла, а верхний, наоборот, охлаждается, как показано на рисунке. — Прим. ред.
1 Многие млекопитающие при тепловой одышке дышат через нос, с закрытым ртом, и только уж в очень сильную жару, когда животные проявляют явные признаки беспокойства, они открывают рот и начинают дышать медленнее, глубже и с большими усилиями.
1 Некоторое изменение частоты дыхания при тепловой одышке допустимо и наблюдается на самом деле. Изменения мышечного тонуса вызывают изменение собственной частоты вязкоупругой колебательной системы.
1 Один из моих сотрудников, Лутц, наблюдая за голубями, подвергнутыми тепловому стрессу, обнаружил у них более сильные колебания частоты дыхания при тепловой одышке, чем сообщает Кроуфорд. Причина расхождения этих данных пока не найдена.
1 Предполагали, что воздушные мешки действуют как набор мехов, у которых чередуются фазы сокращения и растяжения. Например, если задние мешки сокращаются, а передние в это время растягиваются, то воздух мог бы перетекать из одних в другие. Однако, как показали наблюдения, в передних и задних мешках происходит одновременное и почти одинаковое падение давления, что исключает возможность непосредственного перетекания сколько-нибудь значительных масс воздуха из одних мешков в другие.
1 Легкое птицы гораздо менее эластично, чем легкое млекопитающего; будучи извлечено из тела, оно не спадается и сохраняет свою форму.
1 В настоящее время общепризнанно, что у человека потребление кислорода линейно зависит от скорости бега в довольно широком диапазоне скоростей. При очень больших скоростях затраты кислорода возрастают и кривые загибаются вверх. Однако неоднократно высказывались утверждения о том, что эта зависимость не является линейной; и в самом деле, если учитывать точки для очень высокой скорости бега, а также точку для метаболизма в состоянии покоя (которая не совпадает с точкой пересечения с осью у), то получается более сложная, нелинейная зависимость. Тот факт, что точка пересечения с осью у располагается выше точки, соответствующей метаболизму в состоянии покоя, можно приписать «эффекту позы», то есть затрате энергии на поддержание тела в вертикальном положении; например, у неподвижно стоящего человека интенсивность метаболизма близка к точке пересечения кривой бега с осью у. Определение «истинной» величины метаболизма в состоянии покоя (или, как его крайне неудачно называют, «основного обмена») — очень трудная задача, особенно применительно к животным, которых мы не можем «попросить» лечь и расслабиться. Усыпление животного или наркоз не помогают решить проблему, так как при этом интенсивность метаболизма ниже, чем в состоянии покоя.
1 Тепловая одышка дает еще одно преимущество, о котором я уже говорил: она позволяет сохранять высокую температуру на поверхности тела, что автоматически уменьшает перепад между высокой температурой воздуха и температурой кожи животного, уменьшая тем самым создаваемую средой тепловую нагрузку и расход воды. А у животных, у которых терморегуляция достигается с помощью потоотделения, температура поверхности тела относительно невысока (ниже температуры глубоких участков), и это облегчает приток тепла из окружающей среды к коже, повышая тепловую нагрузку и расход воды. Однако мы все еще недостаточно ясно представляем себе относительные биологические достоинства потоотделения и тепловой одышки. По всей вероятности, потоотделение дает какие-то преимущества. Быть может, оно помогает животным с очень редким шерстным покровом или с голой кожей избегать перегревания на солнце?
1 Райнер и Кинан (1967) сообщают, что тунец плавает главным образом при помощи красных мышц, а белые мышцы служат резервом, используемым для коротких вспышек бурной активности.
1 По-видимому, в организме кита поддерживается обычное артериальное давление, необходимое для того, чтобы происходило кровообращение; однако его величина очень мала по сравнению с гидростатическим давлением на большой глубине.
1 При внимательном чтении «Путешествий Гулливера» можно обнаружить некоторое расхождение между допущениями Клейбера и утверждениями Свифта относительно размеров лилипутов. Свифт, естественно, не был знаком с современной физиологией и определял рацион по весу, который он получал возведением линейных размеров в третью степень.
1 Жизненная емкость легких — наибольший объем воздуха, который можно выдохнуть после самого глубокого вдоха. Дыхательный воздух — объем воздуха, поступающего в легкие при обычном вдохе — Прим. ред.
СОДЕРЖАНИЕ
|
| {141} |
Кнут Шмидт — Нильсен
КАК РАБОТАЕТ ОРГАНИЗМ ЖИВОТНОГО
Редактор Р. Дубровская
Художественный редактор Ю. Урманчеев
Технический редактор Л. Бирюкова
Корректор Н. Гиря
Сдано в набор 10/Х 1975 г. Подписано к печати
22/IV 1976 г. Бумага тип. №3 84×1081/32=2,25 бум. л.
7,56 усл. печ. л. Уч.-изд. л. 6,96. Изд. № 12/7609
Цена 33 коп. Зак. № 926
ИЗДАТЕЛЬСТВО «МИР»
Москва, 1-й Рижский пер., 2
Ордена Трудового Красного Знамени
Ленинградская типография № 2 имени
Евгении Соколовой Союзполиграфпрома при
Государственном Комитете Совета Министров
СССР по делам издательств, полиграфии и
книжной торговли
198052, Ленинград, Л-52,
Измайловский проспект, 29
| {142} |
ВНИМАНИЮ ЧИТАТЕЛЕЙ!
В 1977 ГОДУ ИЗДАТЕЛЬСТВО «МИР»
ВЫПУСТИТ КНИГУ
Р. ЗЮСС, В. КИНЦЕЛЬ, ДЖ. СКРИБНЕР
«Рак: эксперименты и гипотезы»
В книге, принадлежащей перу широко известных ученых в области экспериментальной онкологии, дан исчерпывающий обзор различных воззрений на возникновение рака, рассмотрены теории химического канцерогенеза, вирусная теория, генетическая предрасположенность, а также иммунология опухолей и перспективы иммунотерапии рака.
Написана книга легко, превосходно иллюстрирована, снабжена словарем, облегчающим чтение неспециалисту. Ее с большим интересом и пользой прочтут не только медики и биологи, но и все те, кто хотел бы больше узнать об этой актуальнейшей проблеме современной медицины.
| {143} |
|
В 1976 ГОДУ ИЗДАТЕЛЬСТВО «МИР» Б. БОВА «Новая астрономия» Б. БРИНКВОРТ «Солнечная энергия для человека» Э. ДАК «Пластмассы и резинах К. КУЗОВ «Мир без форм» |