
|
П.А.КОРЖУЕВ ЭВОЛЮЦИЯ
ИЗДАТЕЛЬСТВО «НАУКА» Москва 1971 |
| {1} |
|
Освоение космоса ставит перед биологами труднейшую проблему — изучение невесомости. Межпланетные полеты будут проходить в условиях, где практически отсутствует гравитационное поле, т. е. человек и сопровождающие его животные и растения будут находиться в условиях невесомости месяцы и даже годы. Автор книги, известный специалист в области сравнительной физиологии и биохимии, излагает концепцию, согласно которой силы гравитации играли важную роль в эволюции жизни на нашей планете. Исходя из этих положений, он высказывает соображение о вероятных последствиях длительного пребывания человека и животных в состоянии невесомости, подчеркивает, что невесомость — не только космическое явление, но также и земное. Водные организмы (в том числе и гиганты моря — киты) практически находятся в состоянии, близком к невесомости. Книга представит интерес для широкого круга биологов — биохимиков, физиологов, медиков, студентов. Рисунки
|
| {2} |
Предлагаемая вниманию читателей книга профессора П. А. Коржуева касается одного из интереснейших аспектов проблемы эволюции — роли в филогенезе животного мира фактора гравитации — постоянного действия поля земного тяготения на организм. Исходя из широких общебиологических взглядов, автор прослеживает изменения, вызванные переходом живых существ из первичной колыбели жизни — мирового океана — на сушу. Этот переход представлял собой качественный скачок в гипергравитационную среду. Автор справедливо указывает, что переход к иным условиям существования, вызвавший большие морфофизиологические изменения в организмах, не привлек сколько-нибудь широкого внимания физиологов, а стал традиционной монополией морфологии, вследствие чего приобрел известную однобокость. Несколько большее, хотя все же недостаточное освещение получили со стороны физиологов и биохимиков изменения у вторично-водных (перешедших из гипергравитационной в гипогравитационную среду) животных, особенно, млекопитающих.
П. А. Коржуев особенно подробно останавливается в своей книге на изменениях скелета и основного органа кровотворения — костного {3} мозга, связанных с повышенным влиянием гравитации в условиях наземной жизни. Обусловленное этими изменениями усиление энергетического обмена потребовало в свою очередь повышенной доставки кислорода тканям и увеличения массы крови и ее кислородной емкости. Убедительным доказательством высказанной точки зрения служат изменения вторично-водных животных (редукция скелета и костного мозга, уменьшение синтеза гемоглобина в костном мозге и повышение в скелетной мускулатуре).
В связи с освоением космоса вопрос о влиянии длительной невесомости приобретает не только большое теоретическое, но и чисто практическое значение. Однако интерес книги П. А. Коржуева не ограничивается только этим. Благодаря широкой эрудиции автора книгу с интересом и пользой прочтут и широкие круги биологов — эволюционистов и просто любознательных читателей, количество которых так выросло в нашей стране в результате роста культурного уровня народа. В этом одно из величайших завоеваний нашего общественного строя.
Академик В. В. Парин
| {4} |
Эволюция, гравитация, невесомость... Что, казалось бы, общего между этими понятиями? До сих пор они применялись и применяются в разных областях знания.
Эволюция — термин биологический, касающийся закономерностей изменения природы живых организмов на всем протяжении истории жизни на нашей планете. Гравитация — термин физический, характеризующий силы взаимного тяготения масс независимо от того, идет ли речь о земных телах или планетах и звездах, что нашло свое выражение в законе всемирного тяготения, открытым Ньютоном. Невесомость — также термин физический, ставший особенно популярным после первых полетов в космос животных и человека.
Так что же все-таки объединяет эти термины?
Появились факты, свидетельствующие о тесной зависимости, существующей между природой живых организмов и тем гравитационным полем, в котором они живут. Биологи до сих пор рассматривали эволюцию организмов на нашей планете как явление изолированное, самобытное. В первую очередь изучалась хронология появления организмов в ряду современных животных, в меньшей степени — зависимость особенностей структуры и функций организма от особенностей внешней среды.
Однако, несмотря на то, что живые организмы так же, как и неживые тела, подчиняются силам земного притяжения, роль сил гравитации в жизни животных и растений до сих пор находится вне поля зрения биологов. Если характер влияния на живые организмы таких внешних факторов, как тепло, свет, влажность, низкое парциальное давление кислорода и другие, в основном известен, то конкретный характер воздействия сил {5} гравитации остается неизвестным. Совершенно очевидно, что пока не будут выявлены конкретные пути воздействия такого мощного фактора, как силы гравитации, на живые организмы, мы не сможем понять те закономерности, которые определяют эволюцию жизни на нашей планете.
Физика пока не дала ответа на вопрос о том, что такое силы тяготения, или гравитации, какова их природа, как осуществляется взаимодействие между тяготеющими телами.
Силы гравитации резко выделяются в ряду других сил природы. Природа света, электричества, магнетизма известна, их можно генерировать. Все они обладают способностью взаимопревращения, что и используется в современной технике (тепло, например, превращается в электричество, электричество в свет и т. д.). Силы гравитации не могут быть генерированы и пока не поддаются никаким воздействиям со стороны человека. В то же время это самые универсальные силы, проявляющие себя во всех звеньях Вселенной,— от атома до галактик, ибо гравитационное поле создается там, где имеются материальные массы. Все, что имеет массу (а масса присуща любому виду материи), должно испытывать гравитационное воздействие.
Каждая частица материи обладает свойством притяжения к другим материальным частицам. Общая сумма всех притяжений в любой точке Вселенной составляет местную величину гравитации. Различие определяется лишь величиной массы взаимодействующих тел. Если такими телами являются планеты Солнечной системы, то силы гравитации будут определяться размерами планет. Наибольшая сила гравитации на поверхности таких крупнейших планет, как Юпитер, Сатурн, Уран, Нептун (по сравнению с действием сил гравитации на поверхности нашей Земли). Причем, на Юпитере силы гравитации в два с половиной раза выше, чем на Земле, тогда как на Луне они в шесть раз меньше, чем на Земле.
Как известно, Ньютон выразил закон всемирного тяготения простой формулой
|
|||||
где F — |
сила притяжения между двумя телами, |
||||
f — |
коэффициент пропорциональности или гравитационная постоянная, |
||||
m1m2 — |
массы тел, |
||||
r — |
расстояние между телами. |
| {6} |
Суть этого закона состоит в том, что два тела действуют друг на друга с силой, прямо пропорциональной произведению их масс и обратно пропорциональной квадрату расстояния между ними. Примечательно, что обратная пропорциональность квадрату расстояния характерна и для электрических сил. Две электрические частицы, например два электрона, за счет электрического заряда отталкивают друг друга с силой, обратно пропорциональной квадрату расстояния между ними, но благодаря действию сил гравитации притягиваются друг к другу опять-таки с силой, обратно пропорциональной квадрату расстояния. Пока никому не удалось показать, что тяготение и электричество представляют собой проявление одной и той же сущности, ибо в таких масштабах эти две силы несоизмеримы: тяготение относится к электрическому отталкиванию, как единица к числу с 42 нулями.
Однако гравитационные силы становятся грандиозными, когда тела обладают огромной массой, как, например, планеты, звезды. Таким образом, силы гравитации представляют собой дальнодействующие силы.
Физики, характеризуя роль сил гравитации в жизни нашей планеты, подчеркивают, что форма планеты, ее водный и воздушный океан существуют только благодаря силам гравитации: «Волна в море, движение каждой капли воды в питающих это море реках, все течения, все ветры, облака, весь климат планеты определяются игрой двух основных факторов: солнечной деятельности и земного притяжения. Гравитация не только удерживает на Земле людей, животных, воду и воздух, но и сжимает их. Это сжатие у поверхности Земли не так уж велико, но роль его немаловажна.
Корабль плывет по морю. Что мешает ему утонуть — известно всем. Это знаменитая выталкивающая сила Архимеда. А ведь она появляется только потому, что вода сжата тяготением с силой, увеличивающейся с ростом глубины. Внутри космического корабля в полете выталкивающей силы нет, как нет и веса.
Сам земной шар сжат силами тяготения до колоссальных давлений. В центре Земли давление, по-видимому, превышает 3 миллиона атмосфер»1. {7}
О действии сил гравитации можно судить хотя бы по тому, что если бы их не было, то не надо было бы иметь подъёмные краны, лифты в многоэтажных домах, летательные аппараты и многое другое. Все эти сооружения помогают человеку преодолеть действие сил гравитации.
Силы гравитации — фактор огромной мощности, который определяет специфические особенности нашей планеты. Живые организмы, населяющие Землю, не представляют исключения. Они так же, как и тела неживой природы, подвержены воздействию сил гравитации. Мы это воздействие ощущаем на себе буквально на каждом шагу.
Но существует разница в характере воздействия сил гравитации на неживые тела т на живые организмы, специфической особенностью которых является способность к передвижению. Двигаться в гравитационном поле— значит преодолевать его, затрачивать энергию, иметь соответствующие структуры для генерации энергии и для передвижения в гравитационном поле. Именно в этом плане можно ожидать изменений структуры организма животных под влиянием действия сил гравитации.
Эволюция жизни на Земле проходила в гравитационном поле определенной интенсивности, зависящей от размера нашей планеты. И, несомненно, это должно было наложить определенный отпечаток на характер эволюции живых организмов. Именно поэтому переход животных и человека в условия, где гравитационное поле резко ослаблено, то есть в условия невесомости, не может быть безразличным для организма. Рассмотрение этих вопросов и составляет содержание последующих глав.
| {8} |
Глава I
Одна из особенностей Земли как планеты — наличие своеобразной оболочки, именуемой биосферой, которая в свою очередь состоит из трех сфер — атмосферы, гидросферы (моря, океаны) и литосферы (суша). Объединяются они общим наименованием биосферы потому, что являются местом обитания всего многообразия животных и растительных организмов нашей планеты. В настоящее время систематики насчитывают около миллиона видов животных, в том числе около 40 тыс. видов позвоночных животных, 750 тыс. видов насекомых и 500 тыс. видов растений, из них 180 тыс. цветковых и папоротникообразных.
Согласно общепринятым воззрениям все современные животные и растительные организмы существовали на нашей планете не всегда, а появлялись постепенно на протяжении длительной истории нашей планеты, возраст которой определяется в 5—6 миллиардов лет. Таким образом, среди современных животных имеются группы более древние и более молодые (филогенетически более старые и более молодые).
На основании палеонтологических изысканий установлено, однако, что основные типы современных животных уже существовали в кембрии (начало палеозоя), то есть примерно 500 млн. лет назад. В осадочных породах находят остатки червей, иглокожих, ракообразных, моллюсков, хордовых животных, живших в то время.
Появление этих типов животных происходило в определенной последовательности. Древнейшими формами считаются простейшие одноклеточные организмы, которые дали начало многоклеточным, среди которых первыми были губки, затем кишечнополостные, черви, моллюски, ракообразные, иглокожие, хордовые. {9}
Костистые рыбы — филогенетически наиболее молодые среди рыб — появились в силуре (около 400 млн. лет назад), амфибии — в конце девона (около 320 млн. лет назад), рептилии появились в карбоне (около 290 млн. лет назад) и дали начало млекопитающим и птицам.
Первые млекопитающие появились в конце палеозоя, в пермском периоде (около 220 млн. лет назад), птицы — в середине мезозоя — в юрском периоде (около 180 млн. лет). Расцвет млекопитающих и птиц приходится на начало кайнозойской эры (около 65 млн. лет назад). По поводу появления млекопитающих академик А. Н. Северцов писал: «Млекопитающие произошли от весьма примитивных рептилий, по всей вероятности от форм, весьма близких к Териодонтиа, от которых отделились очень давно, по-видимому, еще в течение пермского периода. У нас есть основания предполагать, что в течение всей мезозойской эры потомки этих первичных млекопитающих существовали в виде мелких форм, от которых до нас дошло сравнительно мало остатков. Во всяком случае в течение мезозоя эволюция их шла крайне медленно, и фауна млекопитающих триаса, юры и мела никоим образом не могла конкурировать с крайне богатой и разнообразной рептилийной фауной этих времен. Наоборот, с начала эоцена эта эволюция пошла гораздо более быстрым темпом, так что в продолжение сравнительно весьма короткой третичной эры1 развилось громадное число весьма разнообразных форм млекопитающих»2.
Таким образом, хронология появления представителей основных типов современных животных в основном известна и принимается всеми биологами. Менее ясны причины, приведшие к появлению тех или иных групп животных, и конкретные механизмы изменения природы организма этих животных в процессе эволюции. Не выяснена роль биотических и абиотических факторов в {10} эволюции животных, то есть взаимоотношения организмов между собой и между организмами и средой, окружающей их. По этим вопросам среди биологов еще нет единого мнения.
Следует отметить, что проблемы эволюции животных и растений до недавнего времени привлекали внимание лишь той части биологов, которая изучала строение и форму организмов, то есть морфологов. Другая же, не менее многочисленная часть биологов, изучавшая функции этих организмов,— физиологи и биохимики — уклонялась от разработки проблем эволюции.
В самом деле, все основные положения, касающиеся проблем эволюции животных, хронологии их появления, построения родословного древа, были разработаны морфологами, тогда как физиологи и биохимики в лучшем случае вносили своими исследованиями лишь ясность в некоторые спорные вопросы, например о родстве (близости) тех или иных групп животных, о предках позвоночных животных. Некоторые морфологи считали, что их предками должны быть кольчатые черви. По мнению других, предками позвоночных животных были формы, давшие начало и иглокожим. Следовательно, позвоночные ближе стоят к этой группе животных, чем к кольчатым червям. Биохимики подтвердили это предположение.
Таким образом, необходимость разделения современных животных на первичноротых и вторичноротых обоснована данными. В спорном вопросе о близости осетровых рыб к хрящевым биохимики показали, что осетровые рыбы стоят ближе к костистым, чем к хрящевым. Можно привести другие примеры, которые покажут, что физиологи и биохимики в разработке проблем эволюции стояли на втором плане. Причина, возможно, в том, что изменения формы организмов доступнее анализу.
Совершенно очевидно, что изменение формы организмов в процессе эволюции — лишь последнее звено в сложной цепи процессов, протекающих в организме, начальным звеном которой являются изменения физиолого-биохимических процессов. Несомненно также, что лишь на основе изучения особенностей эволюции формы организмов нельзя понять основных закономерностей эволюции животных. {11}
В сложном комплексе проблем эволюции организмов можно считать решенной, по-видимому, лишь проблему хронологии появления организмов на нашей планете. Тончайшие физиолого-биохимические процессы, обусловившие изменение формы организмов, равно как и факторы, определяющие эти изменения, остаются пока еще недостаточно расшифрованными.
Можно привести целый ряд фактов, касающихся биологии позвоночных животных, установленных морфологами, смысл которых не может быть понят с позиций лишь одной морфологии.
Факт первый. Морфологи не могут объяснить, почему амфибии — первые выходцы на сушу среди позвоночных животных — сбросили чешуйчатый покров, который был у их предков — костистых рыб и превратились в «голых гадов» (по терминологии старых систематиков). А ведь чешуйчатый покров был им очень нужен на суше, может быть даже в большей мере, чем в водных условиях. Такой покров мог удержать воду в организме, ибо потеря воды в условиях суши грозила гибелью.
Факт второй. Вторичноводные млекопитающие приспособились к жизни в воде, видоизменив форму тела на рыбообразную, превратив конечности в лопасти для плавания. Почему разные представители этой группы животных могут нырять на разные сроки: одни на несколько минут, другие — на десятки минут?
Факт третий. Относительный вес скелета новорожденных млекопитающих в два-три раза более тяжелый, чем вес скелета материнского организма. В чем биологический смысл этого явления?
Факт четвертый. Морфолога не могут объяснить, почему рога высокогорных баранов-архаров достигают исключительно мощного развития, часто превышая по весу такую систему организма, как весь скелет в целом. Примечательно, что эти же бараны, обитающие на меньших высотах, обладают менее развитыми рогами.
Здесь приведены факты, на которые можно дать ответ с позиций физиологии и биохимии. Однако есть и такие факты, на которые пока ни морфологи, ни физиологи, ни биохимики не дали ответа. Почему у птиц температура тела более высокая, чем у млекопитающих? Как возникла теплокровность у птиц и млекопитающих? Повысилась ли вначале температура тела, а затем {12} образовался меховой и перьевой покровы для удержания этой высокой температуры тела, или сначала случайно возник меховой и перьевой покров, и только потом благодаря наличию «теплой шубы» температура тела стала повышаться? Совсем не безразлично, какая из двух точек зрения правильная. Но чтобы ответить на эти вопросы, морфологам, физиологам и биохимикам следует работать вместе. Морфология должна быть в теснейшем контакте с науками, изучающими функции организма,— физиологией и биохимией. Без такого сотрудничества с морфологией, без хорошего знания особенностей структуры организма нельзя понять и его функций, то есть тех проблем, над которыми работают физиологи и биохимики. Это очень хорошо понимал выдающийся биолог академик А. Н. Северцов, более полувека назад призывавший физиологов при решении проблем эволюции работать в тесном контакте с морфологами.
Биологи много внимания уделяли изучению роли биотических и абиотических факторов, влияющих на форму организмов. Среди абиотических факторов в центре внимания были температура, свет, влажность, парциальное давление кислорода и другие. В результате накоплено большое количество фактов, свидетельствующих о тесной зависимости формы организмов от особенностей внешней среды. Эта зависимость настолько очевидная, что послужила основанием для формулировки известного принципа о единстве организма и среды, являющегося главным условием успешного изучения биологии современных животных.
Принцип единства организма и среды, впервые сформулированный еще в середине прошлого столетия нашими выдающимися соотечественниками К. Ф. Рулье и И. М. Сеченовым, предполагает детальное изучение всех форм связи, существующих между организмом и средой, его окружающей. Согласно этому принципу в понятие «организм животного» включается не только тело животного, но и среда, окружающая его. «Организм без внешней среды, поддерживающей его существование, невозможен, поэтому в научное определение организма должна {13} входить и среда, влияющая на него. Так как без последней существование организма невозможно, то споры о том, что в жизни важней — среда или самое тело,— не имеют ни малейшего смысла»1. Эти слова принадлежат И. М. Сеченову и были сказаны им более 100 лет назад.
Что же такое внешняя среда?
Обычно под внешней средой биологи понимают в основном комплекс метеорологических и геофизических факторов, которые воздействуют на организм и вызывают в нем те или иные реакции. Существует точка зрения, согласно которой организм в процессе эволюции стремится высвободиться от слишком тесных связей с внешней средой, создать свою собственную внутреннюю автономную среду, характеризующуюся постоянной температурой тела, концентрацией солей, водородных ионов, гемоглобина, сахара и других компонентов крови. Однако эта точка зрения упускает из виду, что «высвобождение организма от слишком тесных связей с внешней средой» (под которым, видимо, подразумевается высокая активность, подвижность организмов) достигается не за счет устранения связей, а, наоборот, как это показано И. П. Павловым, за счет увеличения этих связей, «врастания» организма в среду, совершенствования многочисленных рецепторов, позволяющих животному видеть, слышать, различать запахи на далекие дистанции.
Проблема взаимоотношений организма и внешней среды получила дальнейшее развитие в трудах выдающихся отечественных ученых А. Л. Чижевского и В. И. Вернадского. Около 50 лет тому назад профессор А. Л. Чижевский внес важный вклад в понимание внешней среды, выдвинув положение, согласно которому представление о внешней среде должно быть расширено, ибо внешняя среда включает и мировое пространство, посылающее нам электромагнитные волны разной длины, и потоки частиц, несущих электрический заряд. Другими словами, под внешней средой следует понимать весь окружающий нас мир.
А. Л. Чижевский впервые убедительно показал, что {14} жизнь на Земле теснейшим образом связана с деятельностью Солнца, что периодические явления на Солнце, повторяющиеся через 11-летний период, оказывают огромное влияние на интенсивность размножения и темпы роста организмов нашей планеты. Исходя из этих положений А. Л. Чижевский утверждал, что мир космический и мир земной биосферы связаны воедино, что нельзя органический мир Земли вырывать из природы, отрывать от окружающего мира. Живое связано с окружающей природой, связано с атомами природы всеми атомами своего существа миллионами невидимых связей. Каждый атом живого резонирует на соответствующие колебания атомов природы, сама живая клетка — наиболее чувствительный аппарат, регистрирующий все явления мира и отзывающийся на них соответствующими реакциями организма. Организм неразрывно связан с внешней, так называемой космотеллурической средой. Таким образом, процесс развития органического мира не замкнут, а представляет результат действия земных и космических факторов, из которых вторые являются более важными. Эти мысли А. Л. Чижевским были высказаны еще в 1915 г. в работе «Периодическое влияние Солнца на биосферу Земли».
Представления А. Л. Чижевского о связи биосферы с космосом перекликаются с замечательными идеями академика В. И. Вернадского. Они звучат теперь с особой силой, ибо совершенно очевидно, что характер эволюции живых организмов в значительной мере определялся воздействием факторов внеземного характера. В. И. Вернадский в середине 20-х годов нашего столетия также подчеркивал, что все главные физические проявления Земли как планеты, например ее температурные и физико-химические условия, связаны не только с Солнцем, но и с другими космическими силами, среди которых на первом месте стоят: радиоактивный распад атомов и проникающие космические излучения. Эффект источников космических излучений выше влияния Солнца. Поэтому температурный режим Земли коренным образом иной, чем тот, который возник бы под влиянием Солнца. В. И. Вернадский подчеркивал, что «тепловой режим планеты — не следствие гипотетической остаточной теплоты некогда будто бы расплавленной планеты, молодой Земли, а следствие колоссальной атомной энергии вещества {15} верхней части нашей планеты, вызываемой космическими факторами — радиацией в первую очередь»1.
Касаясь вопроса о причинах, вызывающих распад атомов, В. И. Вернадский писал, что «самопроизвольный» распад вызывают, по-видимому, проникающие космические излучения, «та наибольшая космическая сила, доходящая до Земли, которая вскрывается все больше и больше в своем ведущем значении. Эти лучи идут из областей Космоса, возможно чуждых нашей Галактике, связаны с появлением новых звезд, из областей, где идут самые энергичные процессы — синтез химических элементов из энергии... Солнце по своему значению на нашей планете — по данным геологии — отнюдь не играет той исключительной рели, которую представляет себе планетный астроном. Точно так же и другой вывод планетной астрономии противоречит действительности. Это — образование атмосферы. Земная атмосфера связана не только с всемирным тяготением, т. е. со скоростью отлета, а она биогенного происхождения, связана со скоростью (размножения организмов, т. е. со скоростью биогенной миграции атомов»2.
В биосфере господствуют проявления живого вещества и космических сил в виде радиоактивности и проникающих космических излучений. Поэтому изолировать живое вещество и нас самих от потока космических излучений мы пока не можем. В. И. Вернадский резко критикует распространенный взгляд, согласно которому биосфера — это лишь живые организмы, в ней находящиеся, сторонние самодовлеющие тела, как будто причинно с ней не связанные. В действительности живой организм есть продукт, форма организованности биосферы, а совокупность организмов — живое вещество, ее геологическая функция, геологическая сила исключительного значения.
Считая неверной точку зрения, согласно которой космос и биосфера независимы друг от друга, В. И. Вернадский писал: «Решать биологические вопросы изучением только одного — во многом автономного — организма нельзя. Организм нераздельно связан с механизмом {16} земной коры и должен изучаться в тесной связи с изучением земной коры. Автономный организм вне связи с земной корой реально в природе не существует. Его — для его понимания — надо брать в его среде, в земной коре (правильнее в биосфере). Связь состава организмов с химией земной коры и то огромное — первенствующее — значение, которое имеет живое вещество в механизме земной коры, указывает нам, что разгадка жизни не может быть получена только путем изучения живого организма. Для ее разрешения надо обратиться к его первоисточнику — к земной коре»1.
В то же время жизнь не отделима от космоса, жизнь есть не планетное, а космическое явление. Несомненно, что энергия, придающая биосфере ее обычный облик, имеет космическое происхождение. Она исходит из Солнца в форме лучистой энергии. Но именно живые организмы, совокупность жизни превращают космическую лучистую энергию в земную, химическую, и создают бесконечное разнообразие нашего мира. Это живые организмы, которые своим дыханием, своим питанием, своим метаболизмом, своею смертью и своим разложением, постоянным использованием своего вещества, а главное — длящейся сотни миллионов лет непрерывной сменой поколений, своим рождением и размножением порождают одно из грандиознейших планетных явлений, не существующее нигде, а только в биосфере. Этот великий планетный процесс есть миграция химических элементов в биосфере, непрерывно длящаяся сотни миллионов лет, согласно определенным законам.
В этом плане пророчески звучат слова гениального естествоиспытателя Ж. Кювье: «Жизнь есть более или менее быстрый, более или менее сложный вихрь, направление которого, однако, неизменно; вихрь этот привлекает всегда молекулы одного определенного рода, которые то вступают в его круговорот, то покидают его... Пока движение это длится, тело, в котором оно имеет место, называется живым: оно живет. Когда это движение безвозвратно останавливается, тело умирает... Таким образом, этот жизненный вихрь... соединял на время воедино элементы тела»1. {17}
По этому поводу В. И. Вернадский писал: «Точно и верно определил живой организм Ж. Кювье как непрерывный во время жизни ток — вихрь — атомов, едущий из внешней среды и во внешнюю среду. Организм жив, пока ток атомов продолжается. Он охватывает все вещество организма. Дыханием, питанием, внутренним метаболизмом, размножением создается непрерывно каждым организмом отдельно или всеми ими вместе биогенный ток атомов, строящий и поддерживающий живое вещество»1.
Интересно, что подобную же мысль в 1861 г. высказал и И. М. Сеченов. По его мнению, сущность так называемых растительных актов животной жизни «заключается в непрерывном, всю жизнь продолжающемся, химическом метаморфозе внешнего вещества, поступающего в тело,— метаморфозе, одним из звеньев которого является самое тело»2.
Эта характеристика организма, который нельзя рассматривать как изолированное автономное тело, в отрыве от среды, его породившей, кажется само собой разумеющейся, бесспорной. И тем не менее, не только во времена И. М. Сеченова были ученые, которые хотели видеть в организме автономное тело, имеющее внутри себя все необходимое для существования. Они есть и в наше время. Они утверждают, что принцип единства организма и среды — плод философской неграмотности, что высшие формы материи (т. е. жизни), не могут подчиняться низшим формам материи, что внешняя среда — это всего лишь фон, на котором разыгрывается драма жизни. Руководствуясь подобными установками, нельзя успешно решать актуальные проблемы биологии.
Фундаментальные факты биологии говорят о том, что внешняя среда — это не фон, на котором разыгрывается драма жизни, а неотъемлемая составная часть организма. Практически нельзя провести грань — где кончается организм и где начинается внешняя среда, точно так же как нельзя указать точно, где кончается граница Солнца и где начинается околосолнечное пространство. Физики, например, утверждают, что эту {18} границу Солнца нельзя установить, поскольку она представляет бушующий слой с переменной плотностью раскаленных газов, толщиной во много тысяч километров. А некоторые астрономы утверждают даже, что мы, земляне, живем внутри Солнца, погруженные вместе с Землей в разреженные сферы атмосферы Солнца. И они по-своему правы.
Силам гравитации подвластно движение галактик, звезд и планет. И нет никаких оснований думать, что живые организмы нашей планеты, обитающие в гравитационном поле, находятся вне влияния этих сил. Более того, имеющиеся факты уже сейчас позволяют выдвинуть положение, согласно которому силы гравитации являются одним из самых мощных факторов внешней среды, определяющим форму и размеры организмов животных и растений.
Прошло почти 200 лет со времени открытия закона всемирного тяготения Ньютоном (1687 г.), прежде чем была высказана мысль о важнейшей роли сил гравитации в жизни организмов. Эту мысль впервые высказал в 1882 г. наш знаменитый соотечественник К. Э. Циолковский. В статье «Биология карликов и великанов» при рассмотрении вопроса о соотношении размеров организмов и планеты, на которой обитают эти организмы, К. Э. Циолковский писал: «Снова возникает вопрос: почему в процессе эволюции человек не превратился в лилипута, если так велики выгоды малых размеров?
Во-первых, абсолютная сила органов у больших существ все-таки больше, и в борьбе их с малыми последним приходится плохо. Во-вторых, умственные способности у больших существ все же преобладают. Это прибавляет победные шансы в борьбе. Будь иная сила тяжести на нашей планете, и размер наиболее совершенных людей, как, впрочем, и всех других существ, изменился бы. Например, при уменьшении тяжести а Шесть раз (как на Луне) рост человека мог бы увеличиться в шесть раз, масса в 216 раз, сила мускулов в 35 раз; соответственно увеличился бы и мозг. Такой человек, благодаря силе своих мышц (и обширному уму) {19} оказался бы победителем, несмотря на то, что в борьбе с мертвой природой маленькие люди имели бы больше физических преимуществ»1.
О том, насколько медленно эта мысль прокладывала себе дорогу, можно судить хотя бы по тому, что только в 1897 г. появилась работа английского ученого Крукса, в которой отмечалось, что форма животных, за немногими исключениями, касающимися водных животных, полностью обусловлена силами земного притяжения. Более конкретная характеристика роли сил гравитации была дана в работе немецкого ученого Гертвига, появившейся годом позже, в 1898 г. В ней говорилось о том, что силы тяжести влияют на все тела и все, что каким-либо образом связано в растительном и животном мире с понятием брюшной или спинной стороны, правого или левого бока, носит на себе печать силы тяжести, отражающейся на строении организма, и что особенности строения животных и растений могут быть объяснены влиянием силы тяжести.
В 1917 г., почти два десятилетия спустя, появилась работа английского ученого Томпсона, в которой развивалась мысль, аналогичная ранее высказанной К. Э. Циолковским. Автор утверждал, что размеры животных зависят от размеров нашей планеты и что в случае удвоения ее размеров, т. е. силы притяжения, большинство наземных форм походило бы на коротконогих ископаемых рептилий, а при уменьшении вдвое силы тяжести, наоборот, они стали бы легкими, тонкими, более активными, требовали бы меньше энергии.
Однако эти высказывания не получили достаточного отклика и в дальнейшем появлялись лишь высказывания, касающиеся особенностей структуры тех или иных частей организма в зависимости от сил гравитации. В частности, отмечалось, что особенности кожи дельфинов определяют скорость их передвижения, поскольку дельфин находится в состоянии, близком к невесомости, или что особенности структуры черепа человекообразных обезьян в определенной мере отражают влияние сил гравитации, и даже что билатеральная симметрия рыб определяется уже в момент оплодотворения икры. {20}
Несомненно важное значение для изучения роли сил гравитации в определении структуры организма имеет монография известного морфолога В. Я. Бровара, вышедшая в 1960 г. под названием «Силы тяжести и морфология животных». Эта монография посвящена выявлению зависимости между силами гравитации и строением животного, главным образом на примере изучения сельскохозяйственных животных. В ней содержится ряд положений, представляющих интерес.
В. Я. Бровар исходит из того, что «масса животного как механическая система не изолирована; она зависит от масс других тел и в первую очередь, конечно, от массы нашей планеты. Именно поэтому вес не только скаляр, ко и векторная величина, и вес животного является силой тяготения относительно Земли. Это значит, что всякое живое существо взаимодействует с силовым полем. С момента возникновения и на всем протяжении своего онтогенетического развития, связанного с изменением внешней формы и положения частей внутри целого, организм находится под влиянием тяготения ... Если представить животное перемещенным на другую планету с резко измененными условиями тяготения, то весьма вероятно, что оно не сможет жить в них, именно потому, что животные адаптированы к силам тяжести только данной планеты. Но и в пределах Земли изменение веса животного, например, при переходе из водной среды в наземную, влечет существенные, вплоть до катастрофичных, изменения в ряде функций животного. Даже перемена только направления сил тяжести относительно тела животного при неизменной величине силы веса, и та может оказаться роковой для животного, если оно не приспособлено к подобным изменениям. ... Силы тяжести несомненно влияют на возникновение и изменение многочисленных структурных и функциональных, притом, биологических особенностей любого живого существа нашей планеты. В таком общем утверждении нет в сущности ничего неожиданного или неприемлемого для биологии; в нем давно признана важность влияния силовых полей, в частности тяготения, на растительный и животный организм и накоплен описательный и экспериментальный материал по геотропизмам»1. {21}
В качестве актуальней задачи В. Я. Бровар видит необходимость исследований, вскрывающих «зависимость между строением животного, включая и его форму, и силой земного тяготения». Таким образом, в работе лишь намечаются пути изучения воздействия сил гравитации на организм животных, хотя и не дается конкретное их решение. В. Я. Бровар замечает, что рассматриваемые вопросы представляют значительную степень новизны, а потому при сопоставлении с некоторыми обычными биологическими представлениями и некоторую чуждость, в иных пунктах неясность. Следовательно, за исключением монографии Бровара, в которой содержится экспериментальная попытка выявления зависимости особенностей структуры организма сельскохозяйственных животных от сил гравитации, во всех остальных работах содержатся лишь общие соображения. Однако все они исходят из признания положения, согласно которому силы гравитации представляют собой фактор исключительной важности. Совершенно очевидно, что изучение основных закономерностей эволюции организмов на нашей планете будет невозможно без учета роли сил гравитации.
Однако необходимость выявления роли сил гравитации в эволюции организмов диктуется не только чисто познавательными мотивами. С момента выхода в 1961 г. в космос первого человека, которым был гражданин СССР Юрий Гагарин, перед наукой возникла новая фундаментальная проблема — проблема невесомости. Речь идет не о кратковременной невесомости, измеряемой минутами, часами иди днями, а о длительной невесомости, измеряемой многими месяцами и годами. С такой невесомостью человек будет иметь дело сначала в межпланетных полетах, а затем и в межзвездных.
Как будет реагировать организм человека на длительную невесомость? Безразлична ли длительная невесомость для организма или это полезный фактор, благодаря которому человеку будет легче жить, а может быть это такой же опасный фактор, как космическая радиация?
Биология должна дать ответ на эти вопросы прежде, чем человек отправится на ближайшую планету — Марс или Венеру. Такие полеты будут продолжаться многие месяцы, и космонавты в течение длительного времени будут находиться в состоянии невесомости. {22}
Проблема длительной невесомости — одна из труднейших проблем, связанных с освоением космоса. Трудность состоит прежде всего в том, что в земных условиях пока не представляется возможным воспроизвести состояние длительной невесомости, устранить действие сил гравитации. Именно поэтому надо искать иные пути решения проблемы длительной невесомости.
Однако эта проблема, как и все другие проблемы, связанные с освоением космоса, должна быть решена на Земле. Выявление роли сил гравитации в эволюции жизни на нашей планете — одно из основных условий решения этой проблемы. Исходное положение — принцип единства организма и среды — имеет силу как в земных условиях (при наличии сил гравитации), так и в межпланетных и межзвездных пространствах (где действие этих сил практически отсутствует). Один из благоприятных моментов — то обстоятельство, что на нашей планете имеются зоны, где действие сил гравитации проявляется неодинаково: максимальное действие этих сил характерно для наземных условий, минимальное — для водных.
Говоря о разной интенсивности действия сил гравитации, мы имеем в виду лишь биологический эффект, а не истинно физические моменты, характеризующие гравитационное поле. Физики будут утверждать, что силы гравитации действуют и в водной среде. Они могут сослаться на ряд примеров, подтверждающих правильность своих возражений. Действительно, в батискафе, в подводной лодке люди испытывают действие сил гравитации в такой же мере, как и на суше. Однако речь идет о погружении в воду не батискафа, не подводной лодки, а человека или животных. В этом случае благодаря высокой плотности воды вес организма уменьшается, вследствие чего определенным образом «снимается» эффект действия сил гравитации. Такой же эффект, по-видимому, имеет место и при парящем полете птиц.
Физики могут упрекнуть автора в отождествлении таких понятий, как «вес» и «гравитация». Со своей точки зрения они будут правы. Однако сами же физики заявляют, что для тел, находящихся на земной поверхности и передвигающихся с небольшими скоростями (например для человека), сила веса практически равна гравитационной силе. {23}
Наличие условий, где силы гравитации проявляются «неодинаково», и позволяет выдвинуть положение о возможности изучения роли сил гравитации в эволюции организмов на Земле. Более того, экспериментальные данные, касающиеся особенностей биологии различных групп современных животных, в первую очередь позвоночных животных, уже сейчас позволяют сделать вывод, согласно которому эволюция наземных позвоночных животных представляет собой в основном эволюцию приспособлений, направленных на преодоление сил гравитации.
С этой точки зрения на нашей планете можно выделить два основных этапа эволюции, неодинаковых по своим масштабам, но почти в равной мере оказавших сильное влияние на изменение природы организма — освоение суши наземными животными и заселение океанов вторичноводными животными.
Появление наземных животных — этап эволюции позвоночных животных, который по масштабам изменения природы организма, изменения основных и возникновения новых жизненно важных структур превосходит все другие этапы. В это время шла фундаментальная перестройка организма первых выходцев на сушу в связи с переходом в условия, где действие сил гравитации проявляется в максимальной степени. При анализе этого важнейшего этапа в эволюции позвоночных животных — освоении ими суши — учитывались многие особенности, связанные с дыханием, водным обменом, сменой способов передвижения. Однако специфические особенности перехода из среды, где действие сил гравитации ослаблено, в среду, где действие сил гравитации проявляется в полной мере, оставались без внимания. А ведь если рассматривать эволюцию позвоночных животных в этом плане, то переход от водного к наземному образу жизни следует считать переходом в гипергравитационную среду, точно так же как переход некоторых групп млекопитающих к водному образу жизни (ластоногие, китообразные и другие) можно рассматривать как переход в гипогравитационную среду обитания.
Вот эти естественные эксперименты природы, создавшей условия с «различной» интенсивностью действия сил гравитации и должны послужить материалом для изучения проблемы невесомости, решить которую в наших земных {24} условиях иными путями пока не представляется возможным.
Проблема невесомости связана не только с освоением космоса. Она имеет значение и в наших земных условиях. Ее разработка необходима для понимания закономерностей эволюции некоторых групп современных животных, в частности — вторичноводных, в первую очередь водных млекопитающих, и эволюции всех млекопитающих животных вообще.
Эволюция вторичноводных животных и особенно водных млекопитающих — это прежде всего адаптация к условиям, в которых действие сил гравитации резко ослаблено, а следовательно, близко к состоянию невесомости. С другой стороны, все млекопитающие в период внутриутробного развития находятся также в «гипогравитационных» условиях, то есть, также в состоянии, близком к состоянию невесомости, ибо они находятся во взвешенном состоянии в околоплодных жидкостях. Объяснение этих явлений биологами и клиницистами сегодня мало отличается от взглядов древних греков на роль костного мозга в жизни организма, согласно которым костный мозг — лишь своеобразная подушка для кровеносных сосудов, снабжающих кровью кости скелета. В настоящее время значение околоплодной жидкости также сводится к защите и смягчению возможных механических ударов при резких движениях тела материнского организма. Это, конечно, верно, однако основное значение этих жидкостей состоит в другом.
Истинное биологическое значение адаптации к водным условиям существования как со стороны вторично-водных животных, так и в период внутриутробного развития млекопитающих возможно будет установить лишь с позиций, учитывающих роль сил гравитации в жизни животных вообще и этих групп животных в особенности.
Одна из особенностей наземных условий существования — значительные колебания температуры внешней среды, особенно в некоторых географических зонах. Если в тропиках суточные и сезонные колебания температуры {25} незначительны, то в полярных зонах они достигают максимальных величин. Даже в умеренных зонах сезонные колебания температуры достигают 70° С (+35°, –35°) и более. Естественно, что обитают в таких условиях животные, у которых выработались защитные адаптационные изменения. В этом отношении все современные животные могут быть подразделены на две группы: в первую группу входят животные, температура тела которых определяется температурой внешней среды, во вторую — животные, температура тела которых не зависит от колебаний температуры внешней среды.
Животных с непостоянной температурой тела называют пойкилотермными. К ним относятся вое беспозвоночные и большая часть позвоночных животных — рыбы, амфибии и рептилии. Животных с постоянной температурой тела называют гомойотермными. К ним относятся всего лишь два класса позвоночных животных — птицы и млекопитающие. Для большинства млекопитающих температура тела обычно равна 37°—39° С, для птиц — 40—43° С.
Однако не у всех млекопитающих одинаково хорошо развиты механизмы поддержания постоянной высокой температуры тела. Имеется группа животных, обладающих способностью в теплое время года быть гомойотермными, а в холодное — пойкилотермными. К ним относятся суслики, сурки, хомяки и другие животные. С наступлением осенних холодов они впадают в состояние спячки, которое сопровождается значительным понижением температуры тела (температурный градиент не превышает 2—3°). В это время года животные становятся практически пойкилотермными. Животных, впадающих в спячку на неблагоприятное время года, называют гетеротермными. В частности, в период активности температура тела суслика колеблется от 30 до 39°, ежа — от 31 до 37°, сони — от 31 до 38°, а в период спячки температура тела падает до 2—3° С.
До недавнего времени считалось, что впадают в спячку только млекопитающие. Большинство птиц в неблагоприятное время года, как известно, улетают в теплые края. Оказалось, что некоторые виды птиц также следует отнести к группе гетеротермных животных: в случае неблагоприятных температурных условий у них снижается температура тела. Они впадают в состояние оцепенения {26} при резких суточных колебаниях температуры среды, как это наблюдается у некоторых видов колибри. В частности, у звездчатой колибри при понижении температуры воздуха до +14° температура тела падает до 14,5° С. Наблюдения показывают, что ближайшие родственники колибри — стрижи, также впадают в оцепенение при резком понижении температуры воздуха. У них также в активный период температура тела равна 41°, а в период оцепенения снижается до 20° С.
Среди птиц имеются виды, впадающие в настоящую спячку, длящуюся до двух месяцев. Козодой (Phalaeonoptilus nuttalli), обитающий в западных штатах Америки и Мексике, находится в состоянии спячки до 80 дней. В это время температура его тела снижается до 18° С (а температура воздуха колеблется от 18° до 23°С). Было отмечено даже снижение температуры тела козодоя до 5° С.
В литературе, между прочим, часто допускается путаница в отношении понятия «зимняя спячка». Часто этот термин применяется к истинно пойкилотермным животным, а иногда пишут, что насекомые или рыбы впадают в состояние спячки. Этот термин следует применять лишь к гетеротермным позвоночным животным. В отношении же пойкилотермных животных правильнее говорить о состоянии оцепенения.
Биологическое значение зимней спячки состоит в том, чтобы пережить неблагоприятное время года, когда не только нет возможности добыть необходимый корм, но и температура внешней среды падает значительно ниже 0°С.
Каковы условия и механизмы зимней спячки? Эти вопросы детально рассмотрены в монографии Н. И. Калабухова1. Мы лишь отметим некоторые моменты. Все животные, впадающие в зимнюю спячку, заблаговременно строят специальные гнезда в норах глубоко в земле, до метра и более (рис. 1 и 2). Самые низкие зимние температуры не влияют на температурный режим гнезда: он всегда остается положительным. Накануне залегания в зимнюю спячку, которая у некоторых из этой группы животных длится от 5 до 8 месяцев в году, {27} они накапливают значительные резервы питательных веществ, в первую очередь жира. Количество жира в организме сурков достигает 35% веса тела. Жир расходуется за время зимней спячки.
Таким образом, эта форма адаптации к неблагоприятным условиям почти не изменяет структуры организма, лишь сильно снижает интенсивность жизненных процессов. Может быть наиболее существенным изменениям подвергается функция питания, ибо на время зимней спячки нацело выключается внешнее питание. Естественно ожидать при этом серьезных изменений в функциях желез пищеварительного тракта, в водном обмене и вообще во всем обмене веществ. Так как температура тела животного во время спячки снижается до 2-3° С, то затормаживаются все процессы
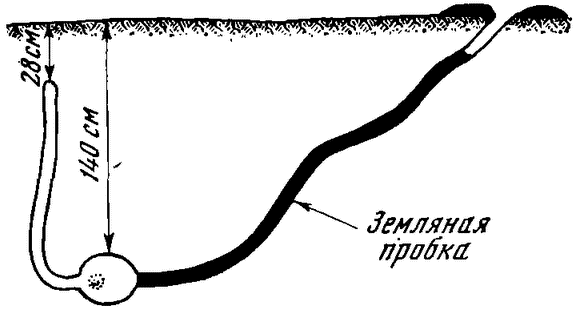 |
Рис. 1. Зимняя нора суслика |
 |
Рис. 2. Суслик в норе во время спячки |
| {28} |
метаболизма. Однако существенных морфологических адаптации мы здесь обнаружить не сможем.
Что касается пойкилотермных животных, то они лишены каких-либо специальных механизмов, предотвращающих влияние температуры внешней среды. Изменения температуры тела пойкилотермных животных, обитающих в воде, пассивно следуют за изменением температуры внешней среды, сохраняя лишь незначительный температурный градиент. Некоторое исключение составляют лишь такие активные рыбы, как тунцы, у которых разница в температуре тела и внешней среды достигает нескольких градусов.
Некоторые из наземных пойкилотермных животных, в частности рептилии, обладают способностью использовать особенности внешней среды для поддержания довольно высокой температуры тела (высокого температурного градиента), а следовательно и высокой интенсивности жизненных процессов. Они выбирают соответствующие условия с иным микроклиматом, обусловленным солнечной радиацией.
Таким образом, у наземных пойкилотермных животных также отсутствуют морфологические изменения, а есть лишь способность регулировать температуру тела путем изменения поведения.
О более серьезных изменениях формы организма в связи с резкими колебаниями температуры внешней среды можно говорить лишь в отношении птиц и млекопитающих. В этом отношении представляют интерес животные Арктики, такие как северный олень, песец, полярная куропатка. Они обитают в исключительно суровых условиях, где температура внешней среды в течение суток и в течение сезона года резко колеблется. Зимой температура в Арктике достигает –70°С, а среднемесячная температура колеблется от –50° до –60°С. Это значит, что в течение многих недель разница в температуре тела животного и внешней среды достигает почти 100°С. В тропиках температурный градиент «тело — воздух» не превышает 10°С, то есть в десять раз меньше.
Чтобы удержать тепло в организме (и сохранить температурный градиент), животные Арктики должны иметь соответствующие морфо-функциональные приспособления. Это в первую очередь шерстный, перьевой покров и подкожный жир. Однако следует иметь в виду, {29} что и шерсть и перья неравномерно покрывают тело, особенно выступающие части тела, такие как конечности, хвост, уши, морда. У арктических животных (канадский и северный олени, горный баран) покровы на ногах в три-пять раз слабее, чем на туловище. Естественно, что эти участки тела значительно теряют тепло. Поэтому для сохранения тепла в организме важное значение имеет поза животного. Так, например, песцы, эскимосские собаки, полярные медведи могут спать на снегу при 50-градусных морозах, свернувшись клубком.
У многих арктических млекопитающих (лемминг, ласка, заяц) конечности гуще покрыты шерстью, чем у тропических. Считается, что теплоизоляция у арктических животных примерно в 10 раз выше, чем у тропических. Однако у многих арктических птиц конечности обнажены, а покрытые пером конечности куропатки и снежной совы, пожалуй, представляют исключение. Вообще же конечности птиц и млекопитающих защищены от холода значительно слабее, чем тело.
Известно, что арктические чайки ходят зимой по снегу при 50-градусных морозах, не обмораживая больших перепонок между пальцами на лапах. Измерение величины теплоотдачи ног чаек в ледяной воде показали весьма низкую величину. Об этом же свидетельствует прямое определение температуры ног, которая оказалась близкой к нулю. Низкая температура конечностей может держаться в течение многих часов, что говорит о способности птиц консервировать тепло в организме. Эта способность свойственна водоплавающим птицам, тюленям, дельфинам, наземным млекопитающим и птицам Арктики.
Способ сохранения тепла в организме при значительном охлаждении конечностей, свойственный многим животным и в более сильной степени развитый у животных Арктики, возможен благодаря особенностям кровоснабжения охлаждаемых частей тела — так называемой противоточной системы кровообращения, а также физико-химическим особенностям жира конечностей. Температура плавления жира, взятого из задней ноги оленя-карибу, понижается от бедра книзу, В частности, костный жир фаланги имеет точку плавления 10—15°, а жир головки бедра 45° С. Жир в подушечках лап не затвердевает при 0°.
| {30} |
Рис. 3. Температура тела и конечностей ибикя |
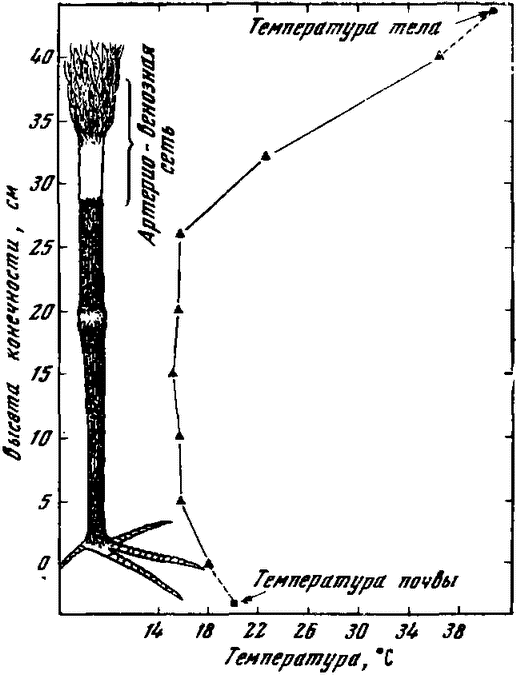 |
Особенностью противоточного кровообращения является своеобразное расположение артериальных и венозных сосудов: крупная центральная артерия оплетается мелкими венами. Эта многоканальцевая артерио-венозная сосудистая сеть играет важную роль в теплообмене и удержании тепла в организме. Благодаря сетевидным сосудистым сплетениям в хвостовых и грудных плавниках китов, тюленей, мышечных впадинах пингвинов температура этих участков тела может быть более низкой, чем температура всего тела. При температуре воды, равной 10°С, температура тела гренландского тюленя равна 10° на поверхности кожи, 22° на глубине 22 мм, 39° на глубине 35 мм. При температуре воды, равной +1°С, температура перепонки между фалангами равна 6—8°С.
Голенастые птицы, такие как журавли, цапли, ибисы, фламинго, имеют тарзиальные артерио-венозные сети (рис. 3). Подобные сосудистые сплетения имеются вокруг конечностей и хвоста у млекопитающих. Благодаря этому температура различных участков конечностей может резко отличаться от температуры их тела.
Может показаться странным, что у животных, обитающих в Арктике, имеются участки тела, недостаточно {31} защищенные от действия низких температур. Целесообразно ли это с биологической точки зрения? Или это одно из проявлений «несовершенства» структуры организма?
Выше отмечалось, что эти участки тела «грают важную роль в тепловом обмене. Животные обычно защищают оголенные части тела от холода, свертываясь клубком. Однако, если животное вынуждено во время бега развивать большие скорости или наступит теплая погода, то в этих условиях возникнет угроза перегрева организма. В таких случаях в оголенных участках тела теплоотдача возрастает в 10—20 раз по сравнению с состоянием покоя. Следовательно, у организма есть возможность даже бороться с перегреванием организма.
Животные Арктики приспособлены для жизни при низких температурах. Хорошим примером может служить северный олень. У него прекрасно развит шерстный покров (даже морда покрывается шестью), и толстый слой подкожного жира. Кроме того, у северного оленя мощно развита третья доля легкого. До сих пор не ясен биологический смысл этой доли легкого. Предполагалось, что поскольку северный олень прекрасный скороход (развивающий скорость до 35 км в час по снежной целине тундры), то у него должен быть очень интенсивный газообмен. Исследования интенсивности газообмена в покое и при быстром беге показали, однако, что потребление кислорода возрастает лишь в семь раз по сравнению с обменом в состоянии покоя. По сравнению с другими млекопитающими это не большое увеличение. Даже у человека во время бега потребление кислорода возрастает в 10—14 раз, а человек не относится к категории лучших скороходов. Другими словами, северный олень мог бы обойтись и двумя долями легкого, как это свойственно другим видам оленей. Тогда зачем нужна третья доля?
Легкие млекопитающих являются не только органом дыхания, но и местом выделения тепла. Если это положение верно, то значение третьей доли легкого северного оленя состоит в выработке дополнительного количества тепловой энергии, столь необходимой в суровых условиях тундры.
Рассмотрение механизмов поддержания постоянной высокой температуры тела, независимой от колебаний {32} температуры внешней среды у высших представителей позвоночных животных, неизбежно вызывает вопрос о том, как возникла теплокровность или гомойотермия в филогенезе позвоночных животных?
По-видимому, проблема происхождения гомойотермии — один из вопросов, успешное решение которых возможно лишь на путях комплексного морфо-физиолого-биохимического исследования. Пока же мыслимы две возможности возникновения гомойотермии. Первая возможность— предки гомойотермных животных сначала приобрели меховой или перьевой покровы, а затем начали накапливать тепло, сохраняя его в организме. Вторая возможность — предки гомойотермных животных уже имели высокую температуру тела, обитая в условиях теплого климата; расселяясь в зоны с более холодным климатом, они сменили чешуйчатый покров на более эффективный шерстный и перьевой. С помощью такого покрова можно было сохранять интенсивность жизненных процессов на прежнем уровне, ибо согласно второму закону термодинамики — никакая энергия на Земле не исчезает, за исключением тепловой, хотя всякая энергия превращается в тепловую.
Второй путь кажется более вероятным. Если стать на противоположную точку зрения, то не ясно, какие же причины вызвали появление шерстного и перьевого покрова? Маловероятно, что это получилось случайно, а затем в результате естественного отбора появилась «шуба» и температура тела увеличилась (у млекопитающих до 37° С, у птиц — до 41—43° С).
Более логична, как нам кажется, вторая возможность, когда в результате адаптации к колеблющимся температурам внешней среды появились шерсть и перо (птицы и млекопитающие произошли, как известно, от разных предков из класса рептилий). В пользу второй возможности появления теплокровности говорят данные температурного режима современных рептилий. Хотя рептилии и относятся к пойкилотермным животным, однако они любят тепло, любят греться на солнце и температура тела у них часто повышается до 40° С (рис. 4). Логично допустить, что у предков млекопитающих и птиц была достаточно высокая температура тела, которую благодаря появлению шерсти и перьев сумели удержать первые млекопитающие и птицы, т. е. они сумели
| {33} |
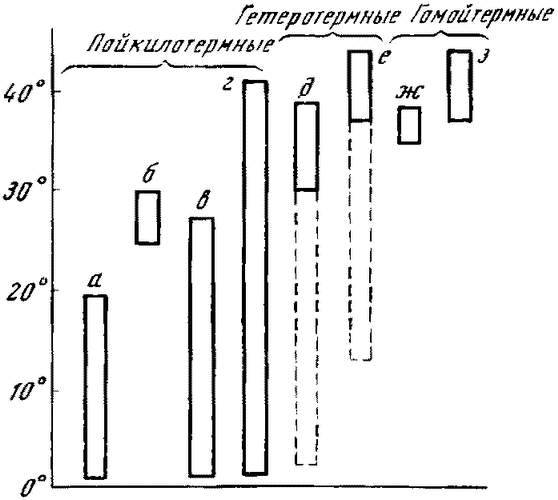 |
Рис. 4. Колебания температуры тела у разных групп позвоночных животных а — рыбы средних широт, б — рыбы тропиков, в — амфибии, г — рептилии, д и ж — млекопитающие, е и з — птицы |
«законсервировать» тот температурный режим, который был характерен для их предков1.
Итак, адаптация к низким температурам у животных Арктики сопряжена с выработкой таких структур, как шерстный и перьевой покров, подкожный жир, третья доля легкого и противоточное кровообращение в некоторых частях тела.
Жизнь в условиях пониженного парциального давления кислорода в высокогорных районах также имеет формы адаптации. В частности, у лам — высокогорных животных, обитающих на высоте 4—5 тыс. м над уровнем моря, гемоглобин крови имеет более высокое сродство к кислороду, чем у равнинных животных. Высокогорные бараны архары и козлы, живущие на высоте 3—5 тыс. м над уровнем моря, обладают мощными рогами. Было много попыток объяснить биологическое значение мощно развитых рогов. Предполагали, что такие мощные рога нужны для прыжков через пропасти, поскольку приземление {34} на рога должно смягчать удар, что такие мощные рога нужны как турнирное оружие в боях за самку. Однако у животных, обитающих на меньших высотах, рога оказывались меньшей мощности. Непонятно, почему на больших высотах для боев за самку надо было иметь рога, вес которых иногда превышал 30 кг и составлял почти 20% веса тела (больше веса скелета).
С нашей точки зрения, столь мощные рога — результат адаптации к пониженному парциальному давлению кислорода. Рога архара покоятся на мощных костных стержнях (рис. 5), которые являются органами, вырабатывающими кровь и гемоглобин, столь необходимый в этих условиях. (Подробно об этом будет сказано в следующих главах.)
Адаптация к условиям засушливого климата, характеризующегося пониженной влажностью воздуха, сопровождается образованием механизмов, предотвращающих потерю воды через поверхность тела. Удержание воды в организме — одно из важнейших условий освоения суши. Для этого у насекомых тело покрылось плотной кутикулой. Рептилии снова обрели чешуйчатый покров, а млекопитающие и птицы — шерстный и перьевой покровы.
Однако наземная среда не представляет собой однотипного комплекса, поскольку имеются зоны более и менее влажные, очень сухие — безводные пустыни, где
 |
|
Рис. 5. Череп горного барана архара а — с рогами, б — с костным стержнем без чехла |
| {35} |
проблема воды приобретает особое значение. Формы адаптации к таким специфическим условиям пустыни различны. Среди животных пустыни известны представители, не пьющие воды вообще. Они пополняют убыль влаги из организма за счет метаболической воды и воды поедаемых растений. В жаркое время года, когда высыхают растения, такие животные, например туркестанский суслик, впадают в летнюю спячку. Другие обитатели пустынь, например верблюд, обладают способностью длительное время обходиться без воды. Несомненно, что эта способность связана с соответствующими адаптационными механизмами удержания воды в организме. Многие из них еще не расшифрованы полностью.
Свет как фактор эволюции играет роль не только в определении суточного и сезонного ритма жизни. Известно, что свет стимулирует созревание половых продуктов у птиц, дает возможность добывать пищу в сумеречное и ночное время. Известно, что некоторые ночные хищники (гремучая змея, филин) обладают способностью видеть инфракрасный свет, что облегчает им нахождение добычи.
Основа реакции на свет — информация фоторецепторов. Они бывают двух сортов: колбочки сетчатки глаза, воспринимающие цветное зрение, и палочки сетчатки — черное и белое.
Подобного рода примеров можно привести много. Однако вывод один: адаптация животных к различным абиотическим факторам (к высокой или низкой температуре, свету, низкому парциальному давлению кислорода, высокой и пониженной влажности и т. д.) отражается лишь на некоторых структурах, существенно не затрагивая морфологию организма в целом.
Адаптация к изменившимся условиям гравитационного поля в наземных условиях вызвала фундаментальные изменения природы организма позвоночных животных (рис. 6). В первую очередь изменился скелет, относительный вес которого значительно увеличился. Скелет позвоночных животных помимо механической нагрузки обрел новую, гемопоэтическую функцию, т. е. стал органом,
| {36} |
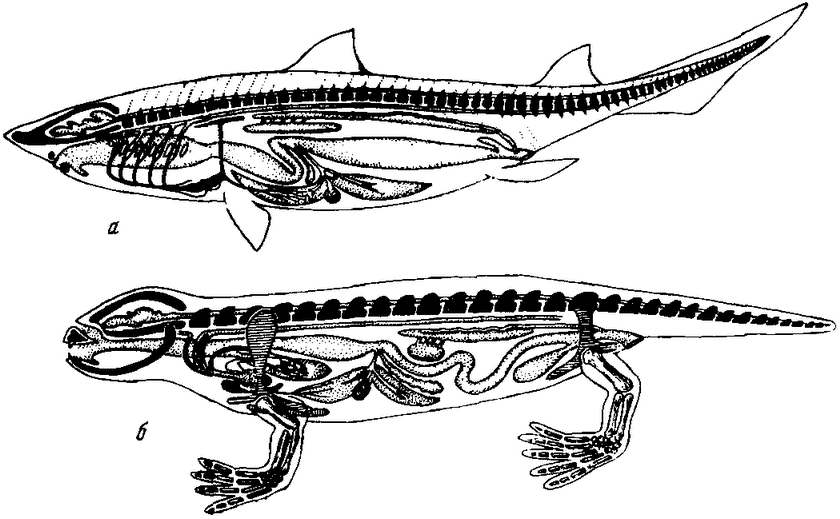 |
Рис. 6. Схема строения тела рыб (а) и амфибий (б) |
вырабатывающим эритроциты и гемоглобин. Появление млекопитающих, характерной особенностью которых является внутриутробный тип развития, по-видимому, обязано именно этой функции скелета. Ведь проблема дополнительных источников кислорода, необходимых материнскому организму и бурно развивающемуся плоду, решается главным образом за счет интенсификации гемопоэтической функции скелета. Организм многих млекопитающих, в том числе и человека, в период беременности претерпевает сильные морфологические сдвиги вследствие изменений некоторых отделов скелета, гемо-поэтическая функция которых в период беременности резко повышается. Количество крови и гемоглобина в крови в материнском организме увеличивается на 25—30%. Все это свидетельствует об интенсификации деятельности скелета как очага синтеза гемоглобина.
Что происходит с дополнительным красным костным мозгом, который функционировал в период беременности? Активная красная кроветворная фракция костного мозга может превратиться в желтую, которая не имеет отношения к кроветворению.
Какова функция этой фракции костного мозга? Можно было бы и не ставить такого вопроса, если бы эта фракция имела ничтожные размеры. Но вес желтого костного мозга у человека составляет от 2,5 до 3% веса {37} тела. Маловероятно, чтобы такая масса ткани, почти сплошь состоящей из жира, оставалась бы простым балластом. Не исключено, что эта фракция желтого костного мозга используется в качестве источника жира для синтеза молока, необходимого для питания новорожденного млекопитающего.
Известно, что скелет имеет самое непосредственное отношение к синтезу молока как источник минеральных компонентов, в первую очередь кальция. Мировая рекордистка — корова Послушница II, давшая за лактацию 16 000 литров молока, погибла в расцвете лет, на 12-м году жизни. Экспертная комиссия не могла установить причин гибели, ибо все внутренние органы были нормальными. И только один член комиссии — В. Я. Бровар обратил внимание на необычную хрупкость и пористость костей скелета. Это был единственный симптом, свидетельствовавший о неблагополучии жизненно важной системы организма коровы. При весьма интенсивном синтезе молока потребность в минеральных компонентах была столь высокой, что мобилизовались все ресурсы организма, в том числе и скелета. В случае с мировой рекордисткой Послушницей II произошла катастрофа вследствие нарушения нормальной деятельности скелета. Это свидетельствует о весьма многогранных функциях скелета, биологическое значение которого еще полностью не раскрыто. Во всяком случае несомненно важное значение скелета в поддержании постоянства внутренней среды организма, или, как теперь принято говорить, гомеостаза.
Особенности гомеостаза как оптимального состояния организма определяются не только положением животного в филогенезе, но и особенностями среды обитания. Каждый организм адаптирован к определенным условиям существования и, следовательно, обладает комплексом структур, поддерживающих это оптимальное состояние. Птицы и млекопитающие как высшие представители позвоночных животных обладают наиболее сложным комплексом структур организма, деятельность которых направлена на поддержание постоянного уровня всех жизненно важных процессов. Именно деятельность этих структур определяет константы, характеризующие гомеостаз того или иного животного: температуру тела, концентрацию водородных ионов, солей крови {38} и лимфы, содержание таких важных компонентов крови, как гемоглобин, сахар и другие. Однако рыбы — низшие представители позвоночных животных — также обладают необходимым комплексом менее сложных структур, деятельность которых определяется своими константами.
По-видимому, все структуры организма принимают участие в поддержании гомеостаза. Так, например, шерстный и перьевой покров поддерживают температуру тела гомойотермных животных, а чешуя рыб — осмотическое давление. Скелет — одна из основных структур организма — и у низших, и у высших представителей позвоночных животных участвует в поддержании определенного уровня солевого состава крови и гемолимфы, а у наземных позвоночных — концентрации гемоглобина крови. Несомненно, что скелет в эволюции позвоночных животных имел важное значение, особенно в период освоения суши, сопряженного с фундаментальной перестройкой организма, и в период появления млекопитающих животных.
С позиций морфологии значение скелета в эволюции позвоночных животных вырисовывалось в виде бледной картины, которая, однако, начинает приобретать более яркие краски, если для выявления его биологического значения привлекаются физиология и биохимия. Поэтому сегодня пророчески звучат слова одного из лучших знатоков скелета сельскохозяйственных животных В. Я. Бровара, который писал: «Интерес зоотехнии к скелету не является, как опасаются некоторые, результатом ее заблуждения относительно потребительской ценности костей. Скелет интересует зоотехнию и не в качестве компонента аппарата движения, как ограниченно трактует его физиология, впрочем, еще не записавшая собственной главы о скелете. Зоотехния рассматривает скелет с гораздо более общей, теоретически и практически несравненно более высокой точки зрения, а именно, как лабильную в онтогенезе сельскохозяйственного животного его часть, обладающую при этом ярко выраженным характером системности»1. {39}
Зоотехники, между прочим, в отличие от морфологов интуитивно чувствовали исключительно важную роль скелета, используя при бонитировке особенности костяка, хотя он и не имеет прямого отношения к тому или иному виду продуктивности животного.
Таким образом, в ряду абиотических факторов роль сил гравитации представляется наиболее фундаментальной, оказывающей огромное влияние на всю природу организма животных, и это влияние по своим масштабам несомненно превосходит воздействие любого другого фактора, о которых речь шла в этой главе.
| {8} |
Глава II
Если бы силы гравитации проявлялись в биосфере Земли одинаково, то наше представление о их роли в эволюции организмов в лучшем случае ограничивалось бы самыми общими сведениями, касающимися формы и размеров животных. Но, как уже говорилось выше, почти 3/4 всей поверхности планеты занимают моря и океаны и 1/4 — суша. Поэтому имеется возможность выявить роль сил гравитации в эволюции жизни на нашей планете путем сопоставления особенностей организмов, обитающих в условиях, где действие сил гравитации ограничено и где действие этих сил проявляется в полной мере, то есть в воде и на суше.
Ботаники лучше зоологов знают воздействие сил гравитации на организм: они воочию наблюдают действие этих сил на растущий организм растения. Если наземное растение в силу случайных причин изменило положение, то растущие побеги примут другое направление, в котором действие сил гравитации будет наименьшим. Известно, что растения на косогорах растут не перпендикулярно поверхности Земли, как это имеет место на ровных местах, а под определенным углом.
Зоологам труднее обнаружить действие сил гравитации на организм животного. Поэтому в зоологических работах можно найти указания лишь самого общего порядка, касающиеся формы организмов и их размеров. Хотя размеры организмов определяются многими факторами, {41} однако бесспорно положение, что животные-гиганты распространены в водной среде, а на суше обитают животные меньших размеров. В связи с этим следует заметить, что бронтозавры и другие гиганты мезозоя, по-видимому, вели земноводный образ жизни, ибо передвижение их по суше очень затруднялось вследствие необходимости преодолевать силы гравитации, действие которых на суше проявляется сильнее, чем в воде. Что касается факторов, определяющих размеры организмов, то представляют интерес соображения, которые высказал академик В. И. Вернадский в статье, посвященной бактериофагу и скорости передачи жизни в биосфере. Характеризуя размеры тел природы, известных нам в настоящее время, он отмечал, что самая далекая звездная система, улавливаемая приборами, отстоит от нас на расстоянии n·1026 см, а самое мелкое тело — ядро атома — протон — достигает размеров n·10–13 см. Таким образом, известные нам тела природы помещаются в пределах 1026 и 10–13 см.
Так как сантиметр — единица произвольная, В. И. Вернадский предложил более мелкую единицу длины, с помощью которой лучше определяется размах наблюдаемых явлений, назвав ее единицей альфа — α. Он считал удобнее принимать альфу равной величине ядра атомов протона — 1·10–13 см, то есть 1·10–6 микромикрона (10–6 мм) или 1·10–5 ангстрема (одна стотысячная ангстрема). Наш мир, выраженный в размерах единицы α, дает величины от 101α до 1039α.
Вернадский считал, что как в космосе, так и на Земле размеры естественных тел не случайны. Согласно предлагаемой шкале предельные размеры живых организмов колеблются в пределах от 107 до 1018α, т. е. от десятков микромикронов до сотен метров, причем 107α, или миллионная часть сантиметра, определяет наименьшую величину живого неделимого.
106α — |
десятимиллионная часть сантиметра — определяет поле действия («размеры») молекул. Это число должно соответствовать максимальному значению величин молекул. |
105α — |
стомиллионная часть сантиметра — определяет поле действия («размеры») атомов. Именно эти величины обнаружены при изучении пространственных решеток кристаллов. |
| {42} |
Очевидно, организмы, тела которых состоят из миллионов молекул и атомов, должны иметь большие размеры, однако их наименьшие размеры приближаются к размерам молекул.
Вернадский считал, что в строении мира размеры 106α—107α не случайны. Величины 105α—106α — это мир атомов и молекул, первостепенное значение которых в строении космоса бесспорно, и 107α — начало мира жизни. Мельчайшие, видимые в микроскоп микроорганизмы достигают нескольких единиц порядка 108α, или десятых долей микрона. Предел зрения ультрамикроскопов 6·107α — шесть сотых микрона. Но есть организмы, размеры которых равны 2·107—3·107α — двум-трем сотым микрона. Здесь жизнь подходит к физическому пределу своего существования. Бактериофаги и являются такими мельчайшими организмами, имея размеры, равные всего двум-трем сотым микрона.
Дыхание этих микроорганизмов отличается от дыхания крупных организмов. Оно осуществляется в поле, регулируемом молекулярными силами: газовый обмен будет идти между телами близкого по величине порядка — между газовыми молекулами и мелкими живыми особями, которые, обладая огромной поверхностью, имеют максимально интенсивное дыхание, а следовательно, и максимальную скорость размножения, возможного для живого вещества. Они же обладают максимальной скоростью заселения планеты, равной скорости распространения звуковых волн (33 000 см в секунду).
Самое замечательное, однако, состоит в том, что организмы, находящиеся на грани живого, сравнимые по величине с молекулами (бактериофаги 107α и 108α), выходят из-под власти поля тяготения, в котором находятся все остальные организмы, и живут в поле действия молекулярных сил. В область проявления сил гравитации входят лишь организмы, имеющие размеры порядка 109α (микроны) и выше, то есть они попадают в наше обычное гравитационное поле, в котором живем и мы, люди.
Однако и в гравитационном поле живое вещество стремится в своем бытии достигнуть всюдности, проникнуть всюду, заполнить среду до конца. Мы не знаем, проявлением чего является это основное свойство живого тела, его стремление заполнить любое пространство — давление жизни. Его проявление и в поле тяготения, и {43} в поле молекулярных сил, по-видимому, свидетельствует о том, что сила, проявлением которой оно служит, выходит за пределы энергетики планеты»1.
Во всяком случае, факт существования форм жизни на нашей планете, не подвластных силам гравитации, примечателен. Правда, основные формы жизни находятся в сфере действия гравитационного поля Земли, и сам по себе этот факт заслуживает внимания именно в плане сравнения с формами жизни, находящимися вне власти сил гравитации.
Для выявления роли сил гравитации в жизни наземных животных, их эволюции нельзя ограничиваться лишь поверхностной характеристикой животных. Необходимо попытаться на основании детального анализа всех жизненно важных структур организма и их функций выявить роль сил гравитации. В этом отношении много может дать анализ особенностей тех структур организма, которые служат для преодоления сил гравитации в наземных условиях. На первом плане, естественно, должно быть изучение костно-мышечной системы, так как скелет и мускулатура — наиболее эффективные механизмы преодоления сил гравитации. Совершенно очевидно, что нормальное функционирование этих структур возможно лишь при соответствующем энергетическом обеспечении, так как преодоление сил гравитации связано со значительными энергетическими тратами.
Что касается особенностей структурных образований, обеспечивающих преодоление сил гравитации в наземных условиях, в первую очередь скелета, то два обстоятельства явились определенным тормозом в выявлений их роли у различных групп животных. Все сравнительно-морфологические исследования носят почти исключительно качественный характер, вследствие чего характеризуются определенной односторонностью. Даже в тех случаях, когда речь идет о редукции частей скелета (наружного или внутреннего), дело сводится лишь к {44} констатации тенденции к редукции, без количественной характеристики явления.
При изучении скелета обращалось внимание лишь на компактное вещество скелета, и совершенно игнорировалась другая органическая составная часть скелета — костный мозг. Более того, когда было установлено, что костный мозг имеет отношение к кроветворению, его стали изучать как самостоятельный орган, имеющий к скелету такое же отношение, как и нервная система. Вследствие этого ученые рассматривали роль скелета лишь в чисто механическом плане, т. е. считали, что скелет выполняет лишь опорно-защитную функцию и является системой рычагов для передвижения.
Таким образом, искусственно разделены две органические составные части скелета, что в свою очередь определило и направление в изучении особенностей скелета. Исследователи интересовались судьбой отдельных частей скелета или отдельных костей, считая, что количественный аспект не имеет значения, ибо с точки зрения механической прочности скелет амфибий уже удовлетворял всем требованиям наземного существования.
Костный мозг интересовал лишь клиницистов, вследствие чего биологические аспекты его формирования не получили должного развития. Ведь костный мозг «запрятан» так же глубоко, как и нервная система. Однако в случае нервной системы ясно, что если нервные клетки располагаются глубоко в недрах организма, то их отростки выходят на поверхность тела, внешнюю или внутреннюю. Следовательно, сигналы, получаемые окончаниями нервных клеток, как-то стимулируют развитие нервной системы, определяют ее эволюцию. Совсем другое дело костный мозг: его клетки находятся или в полостях трубчатых костей или в губчатом веществе костей. Никакой прямой связи с внешней средой нет.
Совершенно непонятно, какие же факторы вызвали к жизни костный мозг и какие факторы определили его формирование и эволюцию в филогенезе позвоночных животных? Такие вопросы и не ставились гистологами и эмбриологами. Изучение сводилось лишь к простой констатации смены очагов кроветворения в онтогенезе тех или иных животных. Даже не было точно установлено, у кого впервые появился костный мозг. По мнению одних исследователей, костный мозг впервые появился у {45} хвостатых амфибий, по мнению других — у бесхвостых амфибий.
Сравнительно-биохимические данные крови, полученные автором книги и его сотрудниками, касались особенностей крови различных групп позвоночных животных. Ниже представлены данные о количестве крови и гемоглобина в организме изученных групп животных:
Количество крови | Количество гемоглобина на кг | |
Рыбы......... | 2,0 | 1,8 |
Амфибии........ | 5,0 | 3,6 |
Рептилии ....... | 5,8 | 4,0 |
Птицы......... | 9,5 | 10,2 |
Млекопитающие .... | 7,5 | 12,1 |
Приведенные данные показывают, что среди позвоночных животных рыбы имеют наименьшее количество крови и гемоглобина, затем следуют амфибии, рептилии. Больше всего крови и гемоглобина имеется у птиц и млекопитающих. У амфибий оказалось почти в два раза больше крови и гемоглобина в пересчете на единицу веса тела, чем у рыб.
Почему у амфибий больше крови и гемоглобина, чем у рыб? В литературе нет ответа на этот вопрос. Первое предположение — у амфибий органы кроветворения более развиты. Известно, что у рыб органами кроветворения являются селезенка, почки или кишечная стенка. Приводим результаты взвешивания этих органов у разных животных в % к весу тела:
Селезенка | Почки | Печень | Костный мозг | |
Северный олень .... | 0,1 | 0,2 | 1,3 | 5,1 |
Морская свинка .... | 0,1 | 1,1 | 4,3 | 1,1 |
Куры (белый леггорн) . . | 0,1 | 0,6 | 1,9 | 2,0 |
Гуси холмогорские .. | 0,1 | 05 | 21 | 2,4 |
Ящерица обыкновенная | 11 | 0,7 | 40 | ? |
Веретеница ...... | 0,4 | 15 | 4 3 | ? |
Лягушка бурая..... | 0,1 | 04 | 3,9 | ? |
Тритон-кристатус ... | 0,1 | 0,1 | 46 | ? |
Карп ......... | 0,3 | 0 7 | 3 6 | Нет |
Сарган......... | 0,1 | 10 | 4 2 | Нет |
Акула катран ..... | 0,3 | 0,7 | 23,1 | Нет |
Скат хвостокол..... | 0,8 | 1.0 | 23,3 | Нет |
Скат шиповатый .... | 0,3 | 0,9 | 56 | Нет |
| {46} |
Данные показывают, что существенной разницы в весе этих органов нет. Тогда было решено обратиться к скелету. Оказалось, что скелет амфибий значительно тяжелее скелета рыб. Скелет бесхвостых амфибий равен 9—12% веса тела, а у рыб в среднем — 7—8% веса тела (таблица 1). Следовательно, переход к наземному образу жизни был сопряжен со значительным увеличением веса скелета.
Поскольку все исследователи признают, что у бесхвостых амфибий в скелете есть костный мозг, можно было предположить, что увеличение в весе скелета происходило не только в связи с усилением механической прочности скелета, но и появлением нового компонента в виде костного мозга.
В основу концепции о гемопоэтической функции скелета1 легло несколько соображений. При переходе от водного к наземному образу жизни появилась потребность в дополнительной энергии для преодоления возросших сил гравитации. Развитие пошло не по пути интенсификации деятельности органов кроветворения (селезенки и почек), а по пути создания нового очага кроветворения в виде скелета. Биологически целесообразно было перемещение очагов кроветворения в те структуры, которые воспринимают всю мощь воздействия сил гравитации в наземных условиях. Такой структурой является в первую очередь скелет. Весь скелет или его отделы (осевой или периферический), воспринимая нагрузку, автоматически могут регулировать деятельность очагов синтеза гемоглобина. Селезенке, почкам, печени труднее регулировать деятельность кроветворных органов. Значит с переходом к наземному образу жизни у позвоночных животных скелет стал выполнять гемопоэтическую функцию, а следовательно, органом кроветворения является не костный мозг, а скелет в целом!
Еще в 1937 г. А. Н. Студитский доказал, что костный мозг и компактное вещество кости происходят из одной
| {47} |
Таблица 1. Весовые особенности скелета позвоночных животных
Животное |
Вес, кг |
Вес скелета, % |
Авторы |
|||
к весу тела |
осевой к весу скелета |
передний пояс к весу скелета |
задний пояс к весу скелета |
|||
Рыбы |
||||||
Форель севанская |
0,3 |
7,1 |
— |
— |
— |
Коржуев, 1956 |
Сарган |
0,4 |
6,4 |
Коржуев, Глазова. 1961 |
|||
Катран |
5,0 |
6,9 |
— |
— |
— |
Те же авторы |
Скат шиповатый |
0,9 |
7,0 |
— |
— |
— |
» » |
Севрюга |
6,0 |
7,7 |
— |
— |
— |
» » |
Амфибии |
||||||
Амблистома |
0,015 |
10,3 |
71,1 |
13,5 |
15,4 |
Коржуев, Акатов, Зубина, 1959 |
Аксолотль |
0,025 |
5,6 |
78,2 |
9,7 |
12,1 |
Те же авторы |
Лягушка травяная |
0,040 |
11,9 |
31,3 |
22,5 |
46,4 |
» » |
Рептилии |
||||||
Агама туркестанская |
0,048 |
13,8 |
66,8 |
15,5 |
17,7 |
Коржуев, Круглова, Свиридова, 1957 |
Степной удав |
0,190 |
14,9 |
100,0 |
Нет |
Нет |
Те же авторы |
Черепаха степная |
0,480 |
43,5* |
31,9 |
39,0 |
29,1 |
» » |
Черепаха болотная |
0,481 |
39,8* |
46,0 |
20,3 |
33,7 |
» » |
Птицы |
||||||
Цесарка |
1,500 |
13,2 |
33,7 |
20,1 |
46,2 |
Коржуев, Корецкая, 1959 |
Утка кряква |
1,328 |
11,2 |
45,4 |
27,3 |
27,3 |
Те же авторы |
Утка пекинская |
2,646 |
9,3 |
49,3 |
27,7 |
23,0 |
» » |
Крачка обыкновенная |
0,102 |
17,9 |
39,7 |
45,6 |
14,7 |
» » |
Млекопитающие |
||||||
Человек |
65,0 |
12,1 |
49,5 |
12,4 |
38,1 |
» » |
Северный олень |
70,5 |
13,4 |
49,2 |
19,6 |
31,4 |
Коржуев. Никольская, 1960 |
Кролик |
4,0 |
6,8 |
49,7 |
16,5 |
33,8 |
Кособутский, 1960 |
Морская свинка |
0,9 |
5,5 |
63,6 |
13,7 |
22,7 |
Коржуев, Науменко, 1962 |
Летучая мышь нетопырь |
0,006 |
17,2 |
34,1 |
53,2 |
12,7 |
Коржуев, Корецкая, 1959 |
Летучая мышь подковонос |
0.009 |
18,8 |
36,9 |
52,7 |
10,3 |
Те же авторы |
* Скелет вместе с панцирем.
| {48} |
и той же остеогенной ткани. Работая с куриным эмбрионом, А. Н. Студитский показал, что первые зародышевые клетки крови, образующие так называемый первичный костный мозг, в котором начинается синтез гемоглобина, появляются только тогда, когда имеется остеогенная ткань, из которой образуется кость. Если нет остеогенных клеток, кровяные островки не образуются. Это значит, что костно-мозговая ткань и костная ткань возникают из одних и тех же клеток — из остеогенной ткани, что костный мозг не посторонний орган для скелета, подобно нервной системе, а органическая составная часть костной ткани.
Если принять эту точку зрения, то становится очевидным, что основным фактором, вызвавшим к жизни костный мозг, являются силы гравитации, ибо он появился у первых выходцев на сушу — хвостатых амфибий. Следовательно, и в дальнейшей эволюции наземных позвоночных животных силы гравитации были тем основным фактором, который определял уровень развития гемопоэтической функции скелета. Это значит, что на скелет нужно смотреть не только как на каркас организма и на систему рычагов для передвижения животного, а как на систему, определяющую благополучие организма. Скелет стал основным очагом синтеза гемоглобина, очагом кроветворения, который в конечном счете решает проблему обеспечения организма кислородом в наземных условиях.
В среде зоотехников до недавнего времени дискутировался вопрос о том, каков должен быть удельный вес скелета у сельскохозяйственных животных? Можно ли снизить относительный вес скелета животных мясного направления и соответственно повысить вес мускулатуры? Постановка такого вопроса вполне правомерна: при весе животного, например коровы, равном 500 кг, более 60 кг веса приходится на кости. Снизить вес костей и повысить вес мускулатуры — перспектива более чем заманчивая.
Делались попытки вывести новые породы тонкокостных животных с более легким скелетом, вес которого составляет 10—12% веса тела, и за счет снижения веса скелета повысить вес наиболее ценной части — мускулатуры. Породу тонкокостных животных пытался вывести английский животновод Беквелл, но его опыт закончился неудачей. {49}
Один из авторитетов в вопросах животноводства-английский ученый Хеммонд писал по этому поводу: «К несчастью, высокая культура разведения крупного рогатого скота привела не только к прорастанию мяса жиром и утолщению мышц, но и к утолщению костей в сравнении с диким необлагороженным скотом, который всегда имеет более тонкую кость. Поэтому следует непременно стремиться при отборе к созданию тонкой кости»1.
Другой авторитет в этих вопросах Дюрст писал: «Много спорят о толщине костей. Швейцарские горные породы скота отличаются толщиной этих костей (речь идет о пястных и плюснёвных костях), что считается скотозаводчиками признаком выносливости породы. И, однако, этот горный скот, будучи импортирован в Северную и Южную Америку, в условиях равнинной пастьбы в обширных степях оказался едва способным сохранить свое существование, не давая соответственной ему хозяйственной производительности, в то время как тонкокостный испанский скот и английский, тоже с более тонкой костью привились в этих условиях хорошо.
...Освещение этому факту дает констатированное в Южной Америке обстоятельство, что в Андах на высоте 3500 метров у скота наблюдается утолщение кожи и костей вместе, что отличает этот скот от низменных пород из льяносов и степей. Следовательно, здесь имеет место влияние климата, обусловливающее конституционные изменения у горного скота. Толстая, крепкая кость у горного скота, так же как более толстая кожа, является приспособлением к изменению внутренней секреции именно в щитовидной железе, и крепкий, выносливый скот в горах нуждается в широких мощных пястных и плюсневых костях. Если они слишком тонки, то это возбуждает подозрение на конституционную аномалию — гипертиресз, располагающую к базедовой болезни, это могут быть молочные коровы с жидким молоком, но отнюдь не хорошие, надежные животные для разведения»2.
Эти верные, по существу, наблюдения животноводов о толстокостности горного скота, объясняются, конечно, {50} неправильно, именно потому, что на скелет смотрят лишь как на механическую систему, определяющую форму организма и образующую систему рычагов для передвижения. Ну и, кроме того, понятия «тонкая» и «толстая» кость — весьма неопределенные. Совершенно очевидно, что «толстокостность» у горного скота есть форма адаптации к высокогорным условиям, где понижено парциальное давление кислорода, и как форма компенсации — утолщение костей. Преодоление сил гравитации в горах требует больших количеств энергии, для чего нужен кислород, гемоглобин. А кислорода в горах мало. Поэтому утолщение костей — реакция организма на эти условия, направленная на увеличение костного мозга, как очага синтеза гемоглобина.
Упрек в недооценке биологического значения скелета следует адресовать не только зоотехникам, среди которых большинство отмечают его большое биологическое значение, но и биологам. Многие биологи-эволюционисты, рассматривая скелет лишь как механическую систему, делали необоснованные выводы об имевшемся в эволюции позвоночных животных переразвитии скелета в целом или его отделов. Американский палеонтолог Ромер считал, что у гигантских рептилий мезозоя имело место переразвитие скелета, который в силу этого был тяжелым бременем для организма. Рассматривая вопрос об особенностях скелета гигантских рептилий, он писал: «Для несения тяжести огромного тела и передачи ее ногам необходима монументальная конструкция позвоночника. Передние позвонки, которым приходилось поддерживать лишь крошечную голову, были малы, но в области туловища телá позвонков становятся все массивнее и массивнее по мере того, как мы переходим через свод спины к области таза. Верхние остистые отростки, короткие впереди, быстро становятся выше и достигают максимальной высоты у вершины свода перед самым тазом ... Но «мертвый вес» самого позвоночника был большим бременем; в соответствии с этим развились приспособления, которые значительно уменьшали вес увеличивающихся позвонков»1. {51}
В руководствах и монографиях по проблемам эволюции причина вымирания гигантского оленя Megaceros eurycerus, жившего в четвертичном периоде, объясняется переразвитием рогов. Среди современных животных мы знаем высокогорных баранов архаров, рога которых также достигают огромной величины. Однако эти животные не вымирают, а наоборот, благодаря своим гигантским рогам, часто весящим больше, чем весь скелет в целом, только и могут обитать в условиях пониженного парциального давления кислорода в горах на высоте 3—5 тыс. метров над уровнем моря. Архары относятся к полорогим животным. У них чехол рога располагается на мощном костном выросте (рис. 5). Мощно развитые рога — это своего рода приспособления к высокогорным условиям существования, при пониженном парциальном давлении кислорода. Мощный костный вырост, на котором располагается чехол, по-видимому, является местом дополнительного образования костного мозга, синтезирующего эритроциты и гемоглобин, который необходим в высокогорных условиях для связывания дополнительного количества кислорода животному. Возможно, что и гигантские олени четвертичного периода обитали в подобных условиях?
Приведенные примеры показывают, что среди биологов распространен весьма типичный подход к оценке скелета, как чисто механической системы, роль который быть лишь каркасом организма и системой рычагов для передвижения, которая в филогенезе позвоночных животных имеет тенденцию к редукции, к снижению собственного веса по отношению к весу дела. Об этом, в частности, писал академик И. И. Шмальгаузен, когда рассматривал вопрос о появлении амфибий: «Первые наземные позвоночные, амфибии, не только не обнаруживают признаков высшей организации по сравнению с кистеперыми и двудышащими рыбами, но даже наоборот, оказываются подвергнутыми процессам редукции в довольно широком масштабе. Особенно сильно редуцировался кожный, но отчасти также и внутренний скелет. Общая жизнедеятельность амфибий не стоит на более высоком уровне, чем у высших рыб. Их активность, пожалуй, даже ниже.
Таким образом, наземные позвоночные произошли от кистеперых рыб путем алломорфоза, а не ароморфоза. Однако вслед за тем облегчение скелета и прогрессивное {52} развитие конечностей привело к увеличению подвижности животного»1.
Об облегчении веса скелета идет речь в учебниках зоологии, когда рассматривается вопрос о полете птиц, для осуществления которого будто бы требуется не только потеря зубов, но и пневматизация костей. Однако все эти весьма ответственные выводы основываются не на точных количественных данных, а на априорных заключениях.
А как обстоит дело в действительности?
В мировой литературе имеется чрезвычайно мало данных по количественной характеристике общего веса скелета и его отделов. Имеется буквально несколько работ по количественной характеристике костного мозга различных животных. Видимо, малая изученность скелета объясняется исключительной трудоемкостью способов очистки скелета. Но основной причиной, конечно, был общепринятый взгляд на скелет, как на чисто механическую систему, которая не представляет интереса с точки зрения выявления эволюционных закономерностей.
Результаты преодоления земного притяжения
Выше были представлены данные о весе скелета и его отделов — периферического и осевого — у некоторых представителей позвоночных животных (табл. 1).
На основании этих данных можно сделать вывод о том, что самый легкий скелет у круглоротых и рыб, и самый тяжелый — у птиц и млекопитающих. Это значит, что в филогенезе позвоночных животных относительный вес скелета повысился (особенно интенсивно в момент перехода от водного к наземному образу жизни).
Однако в процессе эволюции наземных животных также имело место увеличение относительного веса скелета, особенно у птиц и млекопитающих. (Хотя у некоторых представителей этих классов относительный вес скелета такой же, как и у рыб.) Если у крачки вес скелета достигает 18,0% веса тела, у летучих мышей, среди млекопитающих 19,0 (у остроухой ночницы даже 22,0%), то у {53} пекинской утки вес снижается до 9%, а у морской свинки и кролика — до 5,5 и 6,8%. Но эти крайние величины в сторону увеличения и уменьшения относительного веса скелета определяются образом жизни животных и условиями их существования. Морская свинка почти не передвигается, живя всю жизнь в клетке, став классическим объектом лабораторных исследований. У нее самый легкий скелет, более легкий, чем у рыб.
Особый интерес в таблице представляет вес различных отделов скелета у позвоночных животных. Эти данные показывают степень дифференцировки различных отделов скелета в зависимости от положения животного в филогенетическом ряду и, в первую очередь, от образа жизни и условий существования. У первых выходцев на сушу — хвостатых амфибий — на долю осевого скелета у личиночной формы — аксолотля, приходится 80% веса скелета, у наземной взрослой амблистомы — 70%, а на передний и задний пояс и свободные конечности приходится примерно по 15% веса скелета.
У бесхвостых амфибий уже происходит резкое перераспределение веса основных отделов скелета. У травяной лягушки почти половина веса скелета приходится на долю задних конечностей, на осевой скелет — одна треть веса, а на передний пояс конечностей — меньше четверти общего веса скелета. Можно поражаться той колоссальной перестройке организма, которая произошла у бесхвостых амфибий, прежде чем они смогли оторваться от земли и подняться на высоту 30—40 см.
Бесхвостые амфибии были теми первыми существами среди позвоночных животных, которые сделали попытку преодолеть силы земного притяжения в форме прыжка. Однако, какую дорогую цену им пришлось заплатить за эту попытку! 47% веса скелета, 64% веса всей мускулатуры было использовано для совершения прыжка. Среди современных рептилий помимо бегающих форм, подобных ящерице, имеются формы, потерявшие периферический скелет,—змеи и безногие ящерицы. У них 100% веса приходится на долю осевого скелета.
Более разнообразную картину представляет дифференцировка скелета птиц и млекопитающих. У хорошо бегающих птиц, таких, как цесарки, на долю заднего пояса конечностей приходится 46,2% веса скелета, а у хорошо летающей крачки обыкновенной на передний пояс
| {54} |
 |
Рис. 7. Скелет летучей мыши |
конечностей приходится 45,6%. Вес мускулатуры переднего пояса конечностей у крачки составляет 71,1% общего веса мускулатуры, а у цесарки лишь 45%.
У такого прекрасного скорохода, как северный олень, 49,2% веса скелета приходится на долю осевого скелета, 20% на передний и 31,4% на задний пояс конечностей. Наоборот, у малоподвижной морской свинки 63,3% приходится на осевой скелет, 14 и 23% на передний и задний пояса конечностей. И совершенно исключительное распределение веса основных отделов скелета у летающих млекопитающих. У летучей мыши нетопыря-карлика на долю переднего пояса и свободных конечностей приходится 53,2% общего веса скелета, у малого подковоноса 52,7%. Осевой скелет у этих животных составляет 34,1 и 36,9%, задний пояс — 12,7 и 10% (рис. 7). Примечательно, что вес мускулатуры переднего пояса конечностей нетопыря-карлика равен 69,6%, у малого подковоноса 70,8%.
Таким образом, осуществление полета у птиц и млекопитающих потребовало такой же глубочайшей перестройки организма, как и у бесхвостых амфибий, впервые сделавших попытку преодолеть силы гравитации. Примерно половина веса скелета и три четверти веса всей мускулатуры — вот какая цена заплачена за возможность преодолеть силы гравитации на миг и на более длительное парение в воздухе. Нужна была в {55} буквальном смысле слова переплавка организма, фундаментальное перераспределение всех ресурсов организма для решения только одной задачи — вырваться из оков земного тяготения.
Рассмотрение количественных аспектов адаптации наземных позвоночных животных — амфибий, рептилий, птиц и млекопитающих — при освоении суши и воздуха подводит нас к проблеме изменчивости и наследственности, являющейся основной проблемой генетики, вокруг которой идут весьма ожесточенные дискуссии в течение ряда десятилетий. Автор затрагивает эти вопросы только потому, что экспериментальные данные, имеющиеся в его распоряжении и изложенные выше, свидетельствуют о грандиозных масштабах перестройки организма в процесс адаптации к наземным и воздушным условиям существования, то есть касаются проблемы изменчивости В самом деле, если при решении таких важных в жизни организма проблем, как прыжок у бесхвостых амфибий, полет у птиц и млекопитающих, использовались практически все ресурсы организма (более половины веса скелета и до 70% веса всей мускулатуры), то вполне уместен вопрос — а какими темпами шла перестройка организма этих групп животных? В течение какого времени у предков современных лягушек, относящихся к группе бесхвостых амфибий, исчез хвост? В онтогенезе амфибий можно наблюдать процесс исчезновения хвоста у головастиков, а как это происходило в филогенезе?
Ведь основная концепция, выдвинутая еще Дарвином,— представление, согласно которому масштаб этих изменений весьма ограничен Это — мелкие, едва заметные неопределенные изменения, закрепляемые естественным отбором. Трудно себе представить мутацию такого крупного масштаба, чтобы потеря хвоста у предков современных бесхвостых лягушек произошла в один момент. Кроме того, следует учитывать, что потеря хвоста предками амфибий коррелятивно должна вызвать необходимые изменения в других частях организма, в частности, в конечностях А это процесс более сложный Ведь всякие изменения в организме, согласно общепринятым представлениям, носят неопределенный характер. В рассматриваемых же случаях речь идет о весьма глубоких направленных изменениях природы организма, {56} с помощью которых можно было бы осуществить такие сложные действия, как прыжок и полет.
Палеонтологические данные, как отмечалась в главе I, говорят о том, что амфибии появились впервые в конце девона (около 320 млн. лет назад). В мезозое, в меловом периоде (около 130 млн. лет назад) была эпоха расцвета и быстрого вымирания гигантских ящеров — диназавров. И только в кайнозойскую эру (около 60 млн. лет назад) достигли расцвета птицы и млекопитающие. Большие это сроки или малые? Ответить на этот вопрос трудно. Однако, несомненно, что достижение цели было бы маловероятным событием, если бы в основе всего лежал принцип неопределенности и изменения в организме животного шли бы в различных направлениях.
Какой бы точки зрения не придерживался исследователь в вопросе о механизме наследования полезных в данной ситуации свойств, он всегда должен помнить, что организм — это целостная система, любое изменение в которой коррелятивно связано со многими процессами в организме. Именно в силу принципа коррелятивной зависимости, открытого Кювье, трудно допустить возможность появления признаков или свойств, действующих против благополучия такой целостной системы, как животный организм.
В ряде работ, посвященных рассмотрению проблем эволюции, развивается точка зрения, согласно которой, однако, возможно одностороннее развитие тех или иных структур организма вопреки интересам организма как целого. И И. Шмальгаузен по этому поводу писал «Уже при разборе теломорфоза мы встречались с явлениями одностороннего развития органов, но тогда речь шла о специализации, т. е. максимальном приспособлении к ограниченным условиям. Теперь мы имеем в виду развитие признаков и органов, не имеющее явно приспособительного значения или даже, наоборот, являющееся как будто бы помехой для организма (чрезмерно развитые клыки бабируссы или колоссальные рога гигантского оленя четвертичного времени, или, наконец, огромные бивни и общий гигантизм многих слонов)... Нередко намечаются и неблагоприятные моменты Возрастание общей величины тела никогда не бывает пропорциональным, и наиболее дифференцированные части, как {57} например головной мозг, при этом сильно отстают в росте от остального тела». И далее: «По мере возрастания величины тела иногда непропорционально разрастаются различные кожные образования — рога, кожный скелет. Это имело место у цератопсид, особенно у трицератопса — огромного динозавра из верхнего мела. Или кожное окостенение у стенозавра. Эти явления непропорционального разрастания кожных образований обусловлены сохранением закономерностей онтогенетического развития (с его системой корреляций) в быстрых филогенетических преобразованиях»1.
Таким образом, рост бивней у слонов, клыков у саблезубой кошки и бабируссы, рогов у оленей, равно как « разрастания различных кожных образований — рогов, кожного скелета — все эти явления, сопутствующие общему возрастанию величины тела, выделяются своей особенной яркой непропорциональностью. На основании этих положений делается вывод о том, что быстрая эволюция бывает поэтому в некоторых отношениях инадаптивной.
С подобной точкой зрения трудно согласиться потому, что целый ряд примеров можно объяснить совершенно с других позиций. Подобная трактовка, конечно, следствие незнания функций тех или иных структур организма. В качестве иллюстрации этой мысли можно привести пример, использованный Дарвином в «Происхождении видов»: «Обнаженная кожа на голове кондора обыкновенно рассматривается как приспособление для расклевывания падали; это объяснение, может быть и верно, а может быть эта особенность происходит от непосредственного воздействия гниющих веществ, но только с большой осторожностью могли бы мы сделать из этого факта какой-нибудь вывод в отношении нагой кожи на голове всегда опрятно питающегося индюка»2.
Дарвина интересовали причины появления оголенных участков головы и шеи у некоторых видов птиц. Однако понять причины появления этого феномена у хищных (кондор) и всеядных птиц (индейки, казуары) трудно, если не учесть особенностей экологии этих видов птиц. А причина в том, что эти оголенные участки головы и
| {58} |
 |
Рис. 8. Оголенные участки головы и шеи индеек — орган теплообмена |
шеи у индеек, казуаров и кондоров являются органами терморегуляции. Если понаблюдать за индейками, то можно убедиться, что в жаркую погоду у них все выросты на голове и шее наполнены кровью, тогда как в холодную погоду эти образования резко уменьшены в размерах и имеют бледный цвет (рис. 8).
Точно так же и в случае с примерами, иллюстрирующими феномен переразвития. Пока не известны функции тех или иных структур организма, можно говорить о возможности инадаптивной эволюции, В действительности же это результат нашего незнания. Ведь нельзя говорить о рогах самки северного оленя или высокогорного барана архара, что это структуры инадаптивные. Поэтому нет основания говорить, что колоссальные рога гигантского оленя, жившего в четвертичное время, являются инадаптивными. Мы просто не знаем деталей тех экологических условий, в которых приходилось обитать этому гигантскому оленю. Возможно олень обитал в высокогорных условиях или в условиях тундры. Есть основание думать, что огромные рога архара — форма адаптации к высокогорным условиям, специфической особенностью которых является низкое Парциальное давление кислорода. Поэтому нельзя {59} объявлять те или иные структуры организма непропорционально развитыми, инадаптивными. Проще и убедительнее сказать, что мы пока просто не знаем функции того или иного органа, будут ли это рога, клыки, бивни или кожный скелет.
Выше были рассмотрены лишь некоторые количественные аспекты изменения скелета и мускулатуры без учета возможных более глубоких изменений подспудных жизненных процессов. Ведь скелет, как неоднократно подчеркивалось, не просто каркас организма и система рычагов для передвижения животного, хотя важность этих функций никем не отрицается, Скелет позвоночных животных с момента выхода их на сушу обрел новую гемопоэтическую функцию, стал очагом кроветворения — синтеза гемоглобина. К сожалению, количественных определений материального субстрата — костного мозга, где происходит синтез гемоглобина, имеется очень мало. До настоящего времени нет пока ни одной работы по количественному определению костного мозга амфибий и рептилий, в отношении костного мозга птиц и млекопитающих имеется буквально несколько работ.
В таблице 2 представлены данные о количестве костного мозга у взрослых и новорожденных птиц и млекопитающих. Эти данные показывают, что у взрослых млекопитающих больше костного мозга (3,1—5,1%), чем у взрослых птиц (менее 3%). Имеется разница и у новорожденных млекопитающих и птиц, У новорожденного северного олененка количество костного мозга достигает 13,0% веса тела и 58% веса скелета, а у только что выклюнувшихся птенцов — 0,3—0,5% веса тела и 2,5—5,0% веса скелета. Эти различия, несомненно, связаны с типом развития: внутриутробным у млекопитающих и внеутробным у птиц. Птенцы до выклева могут обходиться малым количеством костного мозга потому, что у них в период инкубации кислород проникает через поры скорлупы яйца. У млекопитающих единственный способ получения кислорода — контакт капилляров плаценты с капиллярами стенки матки.
| {60} |
Таблица 2. Количество костного мозга у новорожденных млекопитающих и птиц
Животное |
Вес, кг |
Скелет к весу тела, % |
Костный мозг к весу, % |
Автор |
|
тела |
скелета |
||||
Северный олень: |
|||||
взрослый |
70,0 |
11,4 |
5,1 |
47,0 |
Коржуев, Никольская. 1960 |
новорожденный |
5,9 |
22,6 |
12,8 |
58,3 |
Те же |
Морская свинка: |
|||||
взрослый |
0,9 |
5,6 |
1,0 |
17,0 |
Коржуев, Науменко, 1963 |
новорожденный |
0,1 |
11,5 |
2,8 |
24,5 |
Те же |
Кошка: |
|||||
взрослый |
3,7 |
10,8 |
3,1 |
25,0 |
Коржуев, Счетчикова, 1963 |
новорожденный |
0,11 |
13,2 |
3,5 |
25,1 |
Те же |
Собака: |
|||||
взрослый |
22,0 |
12,0 |
3,1 |
26,0 |
» » |
новорожденный |
0,3 |
18,0 |
4,8 |
29,0 |
» » |
Куры белый леггорн: |
|||||
взрослый |
1,6 |
9,0 |
2,0 |
22,5 |
Коржуев, Модератова. 1963 |
новорожденный |
0,040 |
11,5 |
0,5 |
5,2 |
Те же |
Индейка бронзовая: |
|||||
взрослая |
4,5 |
10,8 |
1,7 |
16,4 |
» » |
новорожденный |
0,060 |
13,5 |
0,3 |
2,4 |
» » |
Гусь холмогорский: |
|||||
взрослый |
5,4 |
9,9 |
1,7 |
17,6 |
» » |
новорожденный |
0,080 |
12,0 |
0,7 |
5,0 |
» » |
Проблема поступления кислорода в организм бурно развивающегося плода в период внутриутробного развития исключительно сложна. До самого последнего времени считалось, что у разных групп млекопитающих плаценты разного типа. Полагали, что капилляры стенки матки отделены от капилляров плода неодинаковым количеством слоев эпителиальных клеток: у одних — пятью слоями клеток, у других — тремя, у некоторых — одним-двумя слоями клеток. Оставалось непонятным, как проникает кислород в организм плода через стенку плаценты путем диффузии, если на его пути столько препятствий? {61}
Недавно, однако, было показано, что дело обстоит несколько иначе. Оказывается, у всех представителей млекопитающих в стенке матки и в плаценте по мере развития постепенно уменьшается количество слоев. К концу внутриутробного периода, когда потребность кислорода особенно велика, капилляры плаценты буквально купаются в материнской крови, что несомненно облегчает диффузию кислорода в организм плода.
Совершенно очевидно, что в последнем случае нужны мощные механизмы по связыванию кислорода для нужд бурно развивающегося плода. Поэтому в организме новорожденного северного оленя в два с лишним раза больше костного мозга по сравнению с материнским организмом, и более половины веса скелета приходится на костный мозг. У птенцов костного мозга меньше, чем в организме матери примерно в четыре-пять раз и, только 2—5% веса скелета птенца приходится на костный мозг. В целом костный мозг новорожденных млекопитающих больше, чем у птенцов примерно в 30—40 раз. Но и у взрослых млекопитающих большее количество костного мозга, чем у взрослых птиц, ибо им приходится для передвижения преодолевать действие сил гравитации, тогда как при полете птиц эти траты значительно меньше.
Такова роль сил гравитации как фактора эволюции позвоночных животных. А как изменился образ жизни беспозвоночных животных в результате перехода на сушу? Следует отметить, что не все группы беспозвоночных животных смогли перебраться на сушу. В частности, не смогли освоить сушу губки, кишечнополостные, иглокожие, ракообразные. Вышли на сушу черви, живущие в основном в почве, моллюски, насекомые.
Черви и моллюски сохранили те же способы передвижения, что и в воде. Однако насекомые использовали рычажный способ передвижения с помощью конечностей, как и позвоночные животные. Кроме того, насекомые освоили воздушную стихию (раньше позвоночных животных). Но насекомые по сравнению с позвоночными животными обладают настолько малыми размерами, что в этом случае можно ожидать иных реакций, чем у позвоночных животных. Размеры тела комаров, мошек так малы, что эти насекомые, по-видимому, находятся в состоянии, близком к состоянию невесомости. Следует {62} учесть, что трахейная система, пронизывающая все тело, значительно облегчает вес тела. Но есть и относительно крупные представители летающих и бегающих насекомых, такие как саранча, стрекозы, тараканы.
К сожалению, мы пока не располагаем данными о весовых соотношениях различных жизненно важных структур организма летающих и бегающих насекомых, чтобы выявить роль сил гравитации в жизни этой многочисленной в видовом отношении группы животных, играющей исключительно важную роль в экономике природы. Эти вопросы еще ждут своего решения.
Однако на основании уже имеющихся данных об особенностях биологии водных и наземных позвоночных животных можно сделать вывод о конкретных путях воздействия сил гравитации на организм позвоночных животных. Биологическое значение зависимости между структурой организма позвоночных животных и мощностью действия сил гравитации трудно переоценить. Исходя из этой зависимости, удалось дать объяснение многим фактам, не имевшим до сих пор рационального объяснения, равно как и выявить роль сил гравитации как одного из самых мощных факторов эволюции животных.
| {63} |
Глава III
Все многообразие фактов, которыми располагает современная биология, свидетельствует о том, что море является колыбелью жизни. В самом деле, растительный мир нашей планеты подразделяется ботаниками на 33 класса, из которых 18 классов — водные формы, а среди обитателей суши насчитывается только 15 классов. В то же время весь животный мир, сгруппированный в 11 типов (простейшие, губки, кишечнополостные, плоские черви, круглые черви, кольчатые черви, моллюски, членистоногие, червеобразные, иглокожие, хордовые), подразделяется на 65 классов, подавляющее большинство которых возникли и живут в море. Достаточно сказать, что только четыре класса позвоночных животных — амфибии, рептилии, птицы и млекопитающие, а из беспозвоночных — насекомые, паукообразные — возникли на суше.
В концепции о возникновении жизни в море, однако, имеется один момент, который требует определенной доработки — это вопрос о происхождении солевого состава Мирового океана. Известная гипотеза Мак-Калума, высказанная им в 1926 г., исходит из предпосылки, суть которой состоит в том, что первобытный океан характеризовался отсутствием солей. Современный солевой состав морской воды обусловлен поступлением солевых компонентов из вод, приносимых реками, и растворимых компонентов побережий и дна Мирового океана. Это {64} постепенный процесс, вследствие чего солевой состав Мирового океана постепенно менялся на протяжении всей истории своего существования. Поскольку история органической жизни на нашей планете связана с определенными геологическими периодами, то в своей гипотезе Мак-Калум пытался объяснить различия в солевом составе полостных жидкостей и крови различных животных возникновением тех или иных групп животных в определенный период, когда солевой состав был иным, чем в настоящее время.
В связи с этим приобретает важность знание длительности различных геологических периодов. Для их определения применяются различные методы, и в зависимости от этого Длительность той или иной эпохи определяется разными отрезками времени. Можно напомнить, что более 100 лет назад лорд Кельвин, исходя из теории охлаждения Земли, рассчитал, что возраст Земли превышает 40 млн. лет. В то время эта цифра поражала воображение, ибо до этого возраст Земли исчислялся тысячелетиями.
К концу 20-х годов нашего столетия появились данные, согласно которым возраст Земли должен быть увеличен до 1 млрд. 700 млн. лет, а к концу 30-х годов стали известны данные, позволяющие увеличить этот возраст уже до 3 млрд. 500 млн. лет. В наши дни геологи считают, что возраст Земли составляет по крайней мере 5 млрд. лет. Как видим, с применением новых, более точных методов исследования Земля становится более древней планетой. Однако биологов и эти сроки длительности истории нашей планеты не удовлетворяют. Весьма оригинальную точку зрения развил акад. Л. А. Зенкевич. Основываясь на показаниях «биологических часов», Л. А. Зенкевич считает, что возраст Земли, определяемый в 4—5 млрд. лет, является заниженным. Он исходит при этом из того факта, что уже в кембрии имелись представители всех основных групп современных животных и что время, отводимое геологами для развития органической жизни, несомненно должно измеряться более крупными масштабами. Срок в 500 млн. лет, принимаемый сейчас для формирования основных типов животных, должен быть увеличен в несколько раз, а возраст нашей планеты, по-видимому, составляет около 10 млрд. лет. Такой срок «подсказывают» биологические часы. {65}
Об этом же свидетельствует и взгляд академика А. П. Виноградова1 на происхождение солевого состава Мирового океана. Вместо гипотезы постепенного засоления Мирового океана за счет солей пресных вод и растворимых компонентов морских побережий и дна Мирового океана А. П. Виноградов высказал идею, согласно которой Мировой океан в момент своего возникновения уже имел тот солевой состав, который характерен для него сегодня. Эта идея интересна тем, что она устраняет основное затруднение, которое стояло перед Мак-Калумом: установление сроков, когда соленость Мирового океана достигла уровня, при котором первые живые существа могли появиться и дать начало многообразию органических форм, характерному для современного Мирового океана. Если же солевой состав Мирового океана был таким, каков он теперь, то жизнь могла зародиться очень давно. Об этом и говорил Л. А. Зенкевич, справедливо считая, что сроки, отводимые для развития органической жизни, слишком коротки.
Эта идея снимает и другую, не менее трудную проблему,— происхождение солевого состава полостных жидкостей, крови, лимфы и гемолимфы современных животных.
Таким образом, утверждение Л. А. Зенкевича получает блестящее подтверждение. Это значит, что первые представители жизни на нашей планете могли появиться не 1,5—2,0 млрд. лет тому назад, а значительно раньше.
На определенном этапе эволюции животного мира, в конце девона (спор лишь о сроках, отделяющих нас от этого времени), появились амфибии — первые выходцы на сушу среди позвоночных животных.
Переход от водного к наземному образу жизни большинством биологов рассматривался как один из важнейших этапов, сопряженный с необходимостью глубокой перестройки организма, в первую очередь органов, ответственных за дыхание и движение. Определенным {66} изменениям подверглись и другие органы, в частности дистантные рецепторы (зрение, слух, обоняние), первоначально адаптированные к водным условиям. У беспозвоночных животных наиболее безболезненный переход из воды на сушу совершили моллюски, такие, например, как представитель брюхоногих — виноградная улитка. Толстая раковина надежно защищает ее от потери воды из организма. Правда, несмотря на это, она обитает в сырых местах и развивает активность только в сырую и дождливую погоду. Дождевые черви тоже в дождливую погоду выходят на поверхность суши, но это лишь эпизоды в их почвенной жизни.
Более драматический характер носило освоение суши насекомыми. Предки насекомых, будучи водными животными, дышали всей поверхностью тела. Но с переходом к наземному образу жизни они столкнулись с серьезной угрозой потери воды из организма. Тело насекомых покрылось толстым слоем плотной кутикулы, которая предохранила организм насекомого от потери воды, но вместе с тем закрыла доступ кислороду в организм через поверхность тела. Это противоречие разрешилось с помощью трахейной системы — сложной системы трубок, по которым кислород доставляется всем клеткам и тканям тела. Эта система снабжения организма насекомых кислородом оказалась, по-видимому, одной из самых эффективных систем, свойственных современным животным. Однако она же стала причиной изменения размеров тела насекомых. Среди насекомых нет гигантов, как это имеет место у позвоночных и моллюсков. Все насекомые обладают весьма малой величиной, потому что пронизанный трахеями организм теряет механическую прочность.
Следует подчеркнуть, что основной материал для суждений о происхождении наземных животных — данные, касающиеся эволюции наиболее детально изученной группы позвоночных животных. Обычно ими оперируют эволюционисты (Северцов, 1939; Шмальгаузен, 1940; Шарский, 1961). Однако обращает на себя внимание то обстоятельство, что появление наземных позвоночных они иногда рассматривали как событие, не более примечательное, чем появление челюстноротых, рептилий или других групп позвоночных животных, то есть событие относительно небольшого масштаба. {67}
Между морфологами и эволюционистами возникали разногласия в оценке этих событий. Так, в частности, А. Н. Северцов рассматривал появление амфибий как ароморфоз, то есть эволюционный процесс, связанный с значительными морфо-физиологическими изменениями, приводящими к новому, более высокому уровню энергетики: «Главным ароморфозмом, вызвавшим усиление общей подвижности тела и подъем жизнедеятельности животных, было превращение их конечностей из плавательных в ползающие; это изменение заключалось в том, что из стенобазальных плавников постепенно образовалась система рычагов наземной конечности, т. е. плечо, предплечье и пясть для передней конечности и бедро, голень и стопа для задней конечности...»1
И. И. Шмальгаузен, наоборот, рассматривал появление амфибий как алломорфоз. Он считал, что общий уровень жизнедеятельности амфибии при этом оставался прежним, если не снизился. Исходя из того, что наземные позвоночные произошли от кистеперых рыб, И. И. Шмальгаузен отмечал, что появление наземных позвоночных животных не сопровождалось общим повышением энергии жизнедеятельности. Фактически здесь был лишь переход из одной среды в другую: из населенной весьма разнообразными позвоночными водной среды в воздушную, ими еще не заселенную.
Биотическая обстановка была тогда на суше более простой, чем в воде. Соответственно первые наземные позвоночные— амфибии — не только не обнаруживали признаков высшей организации по сравнению с кистеперыми и двудышащими рыбами, но даже, наоборот, подверглись процессам редукции в довольно широком масштабе. Особенно сильно редуцировался кожный покров, но отчасти также и внутренний скелет. Общая организация амфибий не на более высоком уровне, чем у высших рыб. Их активность, пожалуй, даже ниже. «Таким образом,— делает вывод И. И. Шмальгаузен,— наземные позвоночные произошли от кистеперых рыб путем алломорфоза, а не ароморфоза»2. {68}
Каждый из ученых по-своему прав, ибо все зависит от того, с какой точки зрения подходить к вопросу. Суть же вопроса в том, что самое существенное в появлении наземных позвоночных животных связано и определяется переходом в «другое» гравитационное поле.
Обращая внимание на детали изменения различных структур организма в результате функционирования их в наземных условиях, морфологи игнорировали то обстоятельство, что наземные и водные условия не равноценны. На первый план они ставили изменения формы организма и в первую очередь рассматривали явные «недоработки» в сосудистой системе и органах аэрации. Но переход от водного к наземному образу жизни был сопряжен с целым рядом весьма драматических ситуаций в связи со сменой жаберного типа дыхания на легочный. При включении легочного кровообращения аэрированная кровь направлялась в желудочек сердца, где смешивалась с венозной кровью. Это обстоятельство могло привести к катастрофическим последствиям в связи с угрозой кислородного голодания. Для предотвращения этих последствий требовался дополнительный орган аэрации. Таким органом стала поверхность тела животных. Для этого понадобилось сбросить чешуйчатый покров, имевшийся у предков амфибий, живших в воде.
Однако чешуйчатый покров в наземных условиях был еще более необходим, чем в водных, в первую очередь для удержания воды в организме. Возникла угроза высыхания. Чтобы предотвратить ее, выходцы на сушу вынуждены были находиться вблизи водоемов. Они были «привязаны» к прибрежной зоне водоемов, то есть практически сушу освоить не смогли. Это сумели сделать лишь рептилии, снова обретшие чешуйчатый покров. Высшие представители рептилий даже достигли полного разделения артериальной крови от венозной. Характерно, что морфологи «не заметили» корреляции между смешением артериальной и венозной крови в желудочке сердца и исчезновением чешуйчатого покрова у амфибий как вынужденной меры, которая дала им возможность {69} перейти к наземному образу жизни. Более того, в усложнении структуры сердца амфибии усматривалось начало прогрессивных изменений сосудистой системы по сравнению с рыбами. В частности, А. Н. Северцов писал: «Прекрасной иллюстрацией филогенетического развития органа, идущего путем ароморфоза, является эволюция сердца позвоночных животных, несомненно повысившая энергию жизнедеятельности активных органов этих животных и обусловившая у них возможность прогрессивной эволюции центральной нервной системы и органов чувств. Эволюция эта, хорошо известная на основании сравнительно-анатомических и эмбриологических данных...1
Ниже (рис. 8) приводится известный схематический рисунок сердца различных представителей позвоночных животных. У рыб венозная кровь проникает в цельное, неподразделенное предсердие, а оттуда в желудочек, из которого течет в аорту и затем в жаберные сосуды, где она окисляется, и окисленная разносится повеем органам. У амфибии предсердие уже отчасти подразделилось. Причем, правая половина осталась венозной, а левая артериальной. В желудочек проникает, таким образом, смешанная кровь. К голове идет преимущественно артериальная часть крови, к вегетативным органам — венозная. У рептилий имеется полное разделение предсердия, но еще не полное — желудочка: здесь левая половина сердца почти целиком артериальная, правая — венозная.
Наконец, у птиц и млекопитающих обе перегородки сердца становятся полыми, так что у них вся левая половина сердца артериальная, вся правая — венозная. У этих животных кровь совсем не смешивается, и их активные органы и органы растительной жизни уже получают вполне окисленную кровь, т. е. у них происходит наиболее интенсивный обмен веществ и наиболее интенсивная жизнедеятельность.
На основе этих данных А. Н. Северцов заключает: «Таким образом, мы видим, что вся эволюция сердца позвоночных животных имеет совершенно определенное направление: она направлена к тому, чтобы доставить всем органам тела животного возможно большее количество совершенно окисленной крови, и это вполне {70} достигается у птиц и млекопитающих. Мы можем легко себе представить, что степень окисления крови при дальнейшем усовершенствовании органов дыхания, т. е. при более совершенном развитии в них капиллярной системы, может еще повышаться, но нам трудно представить себе возможность дальнейшего прогресса в самом сердце и в главнейших сосудах, отходящих от него»1.
Таким образом, согласно Северцову (впрочем, это мнение общепринятое), у низших животных, в частности у рыб, сердце не совершенно, ибо оно состоит всего лишь из двух отделов. Оно становится более совершенным у амфибий и особенно у рептилий, когда появляется третий отдел, — а у некоторых (крокодилов) и четвертый — и достигает предела совершенства у птиц и млекопитающих.
Недостаток этой аргументации в том, что эволюция сердца рассматривается изолированно, вне связи с эволюцией органов дыхания, крови. Совершенно очевидно, что основная причина появления новых отделов сердца не направление эволюции сердца, а смена типов дыхания— жаберного на легочное, обусловленная в свою очередь сменой среды обитания, переходом к наземному образу жизни. Если бы не было смены среды обитания у предков наземных позвоночных животных, то не было бы причин для изменения в строении сердца в этом направлении.
В морфологическом плане освоение суши позвоночными животными привело к возникновению так называемого двойного кровообращения. Причем по-настоящему полное двойное кровообращение появилось лишь у высших рептилий, у птиц и млекопитающих. Что же касается амфибий, то они обладают не полным двойным кровообращением. У них разделены предсердия, но кровь из них эвакуируется в общий желудочек. Во время систолы кровь выбрасывается в артериальный конус, откуда в три главные артерии; в каротидные артерии, несущие кровь к голове, в аорту — во все части тела и в легочно-кожные артерии — к легким и коже (рис. 9).
Еще в ранних работах высказывались предположения, что, несмотря на несовершенное разделение следует ожидать приспособлений, направленных на реализацию {71} совершенного двойного кровообращения. Однако многие считали, что в желудочке амфибий имеет место полное смешение артериальной и венозной крови (рис. 10). Было проведено количественное определение двойного кровообращения у амфибий, у зеленой и травяной лягушек. Применялся метод перфузии раствором Тироде двух сортов — прозрачного, бесцветного и окрашенного краской синей Эванса, имитирующих артериальную и венозную кровь. На основании колориметрических определений степени разбавления перфузата от всех главных артерий судили о функциональном разделении артериальной и венозной крови.
 |
 |
Рис. 9 |
Рис. 10 |
Рис. 9. Схема строения сердца А — у рыб: 1 — ток крови к жабрам, 2 — аорта, 3 — артериальный конус, 4 — желудочек, 5 — клапаны, 6 — предсердие, 7 — клапаны, 8 — венозный синус, 9 — вены от органов тела; Б — у теплокровных животных (птиц и млекопитающих): 1 — легочные артерии, 2 — аорта, 3 — полулунные клапаны, 4 — желудочки, 5 — трехстворчатый клапан, 6 — предсердие, 7 — легочная вена, 8 — вены от органов тела Рис. 10. Анатомическое расположение кровеносных сосудов лягушки, через которые производилась перфузия 1 — нижняя полая вена, 2 — желчный пузырь, 3 — печень, 4 — артериальный ствол, 5 — легочная артерия, 6 — кожная артерия, 7 — аорта, 8 — сонная артерия внутренняя, 9 — сонная артерия внешняя, 10 — луковица сонной артерии, 11 — легочная артерия, 12 — легочная вена, 13, 14 — ветви брюшной вены, 15 — воротная вена печени |
|
| {72} |
 |
Рис. 11. Схема распределения артериальной и венозной крови в сердце и крупных сосудах сердца лягушки А — по гипотезе разделения крови: 1 — желудочек, 2 — спиральная складка, 3 — правое предсердие, 4 — легочно-кожная ветвь, 5 — аорта, 6 — сонная артерия, 7 — отверстие артериального конуса к кожно-легочной ветви, 5 — левое предсердие Б — по гипотезе равномерного смешивания крови: 1 — желудочек, 2 — спиральный клапан, 3 — правое предсердие. 4 — начало легочно-кожной ветви, 5 — сонная артерия, 6 — аорта, 7 — легочно-кожная ветвь, 8 — левое предсердие |
Соотношение между сонной артерией (22,5%), кожно-легочной артерией (34%) и аортой (43,5%) выражается как 2:3:4. Опыты показали, что внутри желудочка имеет место частичное разделение артериальной и венозной крови, приводящее к функциональному разделению между сонной и легочно-кожной артериями. Причем в сонных артериях протекает более артериализованная кровь (рис. 11). В нормальных условиях разделение достигает 50—70% в сонных и легочно-кожных артериях и 35% — в аорте.
При любом типе дыхания имеют место значительные колебания величины сердца, что обусловлено образом жизни, условиями существования. Ниже представлены данные о весе сердца некоторых представителей позвоночных животных.
| {73} |
Животные |
Вес тела, г |
Вес сердца, г |
Сердечный индекс % |
Круглоротые |
|||
Минога....... |
298 |
0,825 |
2,77 |
Хрящевые |
|||
Акула ....... |
2810 |
2,935 |
1,04 |
Скат........ |
1100 |
1,310 |
1,19 |
Костистые |
|||
Карп........ |
1840 |
2,27 |
1,23 |
Пеламида...... |
1600 |
2,9 |
3,05 |
Камбала ...... |
142,6 |
0,065 |
0,45 |
Амфибии хвостатые |
|||
Тритон....... |
6,73 |
0,012 |
1,83 |
Бесхвостые |
|||
Жаба........ |
44,9 |
0,144 |
3,20 |
Лягушка...... |
33,4 |
0,093 |
2,78 |
Квакша....... |
4,7 |
0,031 |
6,52 |
Рептилии |
|||
Медяница...... |
16,56 |
0,027 |
1,61 |
Ящерица ...... |
11,64 |
0,026 |
2,24 |
Уж......... |
91,6 |
0,294 |
3,20 |
Птицы |
|||
Фазан ....... |
1337,3 |
5,502 |
4,19 |
Филин ....... |
1875 |
8,810 |
4,70 |
Чайка ....... |
1336 |
13,30 |
9,98 |
Сорока....... |
231,5 |
2,46 |
10,61 |
Страус....... |
7100 |
76,68 |
10,80 |
Голубь....... |
443,4 |
6,14 |
13,85 |
Жаворонок..... |
35,7 |
0,615 |
17,23 |
Пеночка ...... |
9,35 |
0,182 |
19,46 |
Колибри ...... |
4,02 |
0,1147 |
28,50 |
Млекопитающие |
|||
Кролик домашний . . |
1987 |
5,35 |
2,70 |
Заяц русак..... |
3741 |
37,90 |
8.90 |
Еж......... |
886,7 |
3,36 |
3,79 |
Крот........ |
68,2 |
0,397 |
5,82 |
Мышь ....... |
18,12 |
0,127 |
7,04 |
Выдра ....... |
5 005 |
40,1 |
8,02 |
Летучая мышь ... |
3,73 |
0,054 |
14,36 |
Кабан ....... |
93 000 |
365,9 |
3,93 |
Архар ....... |
60 000 |
552 |
9,20 |
Овца........ |
42 800 |
194 |
4,50 |
Слон........ . |
3 000 000 |
14 100 |
4,70 |
Кит ........ |
100 000 000 |
400 000 |
4,00 |
Дельфин-белобочка |
54 200 |
262 |
4,80 |
Тюлень |
17 000 |
170 |
10,00 |
| {74} |
Следует заметить, что, несмотря на значительный Диапазон колебаний величины сердца у позвоночных животных от 0,45% до 28,50%, все же рыбы в целом обладают значительно меньшей величиной сердца, чем любая другая группа наземных позвоночных животных, что хороша видно из приводимых ниже средних цифр:
Круглоротые...... | 2,77 | Амфибии бесхвостые .... | 4,16 |
Хрящевые....... | 0 96 | Рептилии......... | 2 35 |
Костистые....... | 1,17 | Птицы .......... | 13,22 |
Амфибии хвостатые ... | 1 83 | Млекопитающие...... | 7,56 |
Это обстоятельство несомненно связано с тем, что размеры сердца определяются в значительной мере той массой крови, которая содержится сосудистой системой. Количество же крови в организме определяется энергетическими затратами, которые характерны для тех или иных животных, ибо важнейшим компонентом крови является гемоглобин. Обращает на себя внимание тот факт, что сердечный индекс рыб, выражаемый в промилле (%о), в среднем для рыб равен единице, у хвостатых амфибий почти удваивается, а у бесхвостовых амфибий составляет в среднем 4,16%0.
Рептилии (как бегающие, так и ползающие) обладают сердечным (Индексом, близким к сердечному индексу хвостатых амфибий. Несколько увеличивается сердечный индекс птиц — в среднем он равен 13,22%, хотя колебания у разных видов птиц довольно значительны. У фазанов он снижается до 3,54%, а у колибри поднимается до 28,50%. Это — высший сердечный индекс, известный среди современных животных. У млекопитающих средняя величина индекса равна 7,56% (с колебаниями от 2,70 у кролика до 14,36% У летучей мыши).
Примечательно, что разница в величине сердечного индекса рыб и наземных позвоночных, несмотря на отдельные исключения, подтверждает основной тезис, согласно которому энергетические траты организма в водных условиях меньше, чем в наземных. В то же время сердечный индекс млекопитающих и птиц в среднем в 7—13 раз больше, чем у рыб, у лучших летунов среди птиц, таких, как колибри, в 28 раз, а у летучих мышей — в 14 раз. Это значит, что для преодоления сил гравитации {75} на Земле требуются большие траты энергии, однако для отрыва от Земли и полета в воздухе нужны еще дополнительные усилия.
Величина сердца у разных представителей позвоночных животных взаимосвязана с весовыми особенностями скелета и мускулатуры, точнее тех отделов скелета и мускулатуры, которые берут на себя основную нагрузку по преодолению сил гравитации. У амфибий — это скелет заднего пояса конечностей и мышц конечностей, у летающих млекопитающих и птиц — скелет переднего пояса конечностей и мышц, его обслуживающих.
Таким образом, условием отрыва от земли для прыжка и полета в воздухе являются использование не только 50% веса скелета и 70% всех мышц, но и увеличение размеров сердца в 14 и 28 раз (по сравнению с сердцем рыб).
Следовательно, не будет преувеличением сказать, что все структуры организма, определяющие уровень энергетики животного, перестраиваются для достижения одной цели — преодоления сил гравитации. Это и скелет, и мускулатура, и кровь, и гемоглобин, и сосудистая система. Одной из основных характеристик перестройки является сердечный индекс. Таким образом «прогресс в самом сердце» направлен не на создание «перегородок», а на удовлетворение потребностей организма в самых различных условиях, и это хорошо видно при анализе особенностей сердечного индекса у разных представителей позвоночных животных. Сложный процесс преобразования сердца наземных позвоночных животных завершился полным разделением артериальной и венозной крови.
Изменения функций у первых выходцев на сушу изучены в меньшей степени. Сравнительный анализ энергетических процессов в новых и исходных условиях выявил функциональную мощь органов аэрации — легких и кожи. Оказалось, что у амфибий через поверхность тела осуществляется примерно половина общего газообмена, а у некоторых представителей (безлегочные саламандры) и все 100% газообмена.
Сравнительных данных по энергетическим тратам организма рыб и амфибий имеется мало. Впрочем, их едва ли можно считать убедительными. Более убедительны данные по интенсивности дыхания представителей одного {76} и того же вида, начиная с ранних стадий развития, протекающих в воде, и кончая взрослыми формами, ставшими наземными. Вообще амфибии в этом отношении представляют особый интерес как формы, повторяющие в своем онтогенезе путь, связанный с освоением суши, с переходом от водного к наземному образу жизни.
Имеющиеся пока весьма немногочисленные данные свидетельствуют о том, что в онтогенезе хвостатых амфибий, в частности у аксолотля (водной формы) и амблистомы (взрослой наземной формы) уровень энергетических затрат при переходе от водного к наземному образу жизни возрастает примерно вдвое. Как уже ранее отмечалось, у взрослых амфибий значительно возрастает количество крови в организме и концентрация гемоглобина в крови, а следовательно, и общее количество гемоглобина.
Для сравнения количества гемоглобина в организме различных представителей позвоночных животных как водных, так и наземных требуется особый подход. Размеры тела позвоночных животных колеблются в весьма широких пределах и имеют вес от 2,5—3,0 г (у землеройки) до 150 т (у синего кита), то есть различаются по весу в десятки миллионов раз. В связи с этим требуются величины, дающие возможность сопоставлять особенности разных представителей позвоночных, полученные путем пересчета количества гемоглобина на единицу веса тела.
Исходя из того, что количество гемоглобина в организме косвенно отражает уровень интенсивности метаболизма различных представителей позвоночных животных, в основу оценки масштаба энергетических трат были положены данные по количеству гемоглобина, приходящегося на килограмм веса тела, или обеспеченность организма гемоглобином. Таким образом появляется возможность судить об эффективности скелета как гемопоэтической системы у различных представителей позвоночных животных. Это, несомненно, должно найти отражение в данных по количественной характеристике обеспеченности организма гемоглобином у амфибий, которые {77} первыми вышли на сушу, и у других представителей позвоночных, уже освоивших новую среду обитания — сушу.
Сравнение этих данных показывает эффективность нового очага кроветворения, для которого природа использовала одну из основных структур организма — скелет, отказавшись от интенсификации деятельности прежних очагов синтеза гемоглобина — селезенки, почек, кишечной стенки, которые функционируют в водной среде у предков амфибий — рыб.
Как уже говорилось выше, выход на сушу сопряжен со значительными энергетическими тратами, необходимыми для преодоления резко возросшего действия сил гравитации. В воде организм рыб взвешен, поэтому энергия необходима лишь для передвижения. В наземных условиях для передвижения необходимы специальные органы, которые поддерживают тело на весу и позволяют преодолевать силы гравитации. Они возникли в скелете в виде конечностей. Плотность воздуха почти в 800 раз меньше плотности воды, поэтому действие сил гравитации в воде «снимается». В наземных условиях организм животного попадает в «гипергравитационные» условия, если брать за исходный уровень биологический эффект действия сил гравитации в воде. Совершенно очевидно, что вести активный образ жизни в этом «новом» гравитационном поле возможно лишь при усилении энергетических затрат. А это означает усиление расхода кислорода, что в свою очередь должно стимулировать синтез гемоглобина, повышение его концентрации в крови, а возможно и увеличение количества крови в организме.
У первых выходцев на сушу, как и у наземных животных, появившихся позднее, не произошла интенсификация деятельности очагов синтеза гемоглобина водного периода жизни (селезенка, почки, кишечная стенка), а появился новый очаг синтеза гемоглобина. Им стал скелет. Почему природа отказалась от интенсификации деятельности прежних очагов синтеза гемоглобина, почему она создала новый очаг, использовав для этого скелет? По-видимому, прежние очаги синтеза гемоглобина не могли удовлетворить возросшие потребности организма в гемоглобине.
В главе II приведены данные по весу селезенки, почек, печени и костного мозга водных и наземных животных. {78} Они показывают, что масса костного мозга превышает массу селезенки и почек в десятки и сотни раз. Не исключено, что одной из причин смены очагов синтеза гемоглобина была невозможность увеличения массы селезенки и почек. В этом случае они заняли бы большой объем, создавая трудности для нормального функционирования остальных органов брюшной полости. Но вероятнее, что основной причиной передислокации очагов синтеза гемоглобина при переходе к наземному образу жизни была необходимость в более оперативном контроле за деятельностью этих структур. Ведь на скелет ложится вся тяжесть пребывания в наземных условиях, с его помощью животное поддерживает тело на весу и, кроме того, преодолевает силы гравитации при передвижении.
Скелет в целом и различные его отделы (периферический и осевой) испытывают всю тяжесть нагрузки при стоянии и передвижении. Поэтому логично допустить, что величина нагрузки на скелет является тем стимулятором, который действует автоматически, регулируя интенсивность синтеза гемоглобина. Деятельность очагов синтеза гемоглобина может более эффективно регулироваться, если они находятся в скелете. В этом случае они буквально «ощущают» тяжесть нагрузки. Когда органы кроветворения расположены в брюшной полости, которая почти не имеет отношения к преодолению сил гравитации, они могут удовлетворить лишь животных, обитающих в воде, в состоянии, близком к невесомости.
Если верны эти предположения, можно ожидать, что мощность очагов синтеза гемоглобина в разных отделах скелета будет пропорциональна нагрузке, которую он испытает при преодолении сил гравитации организмом. Следовательно, можно будет предсказать характер размещения костного мозга в различных отделах скелета животного.
К сожалению, в мировой литературе имеется очень мало данных по количественной характеристике костного мозга у человека и животных. Имеется всего несколько работ, касающихся количественной характеристики костного мозга человека, собаки, кролика, морской свинки, некоторых видов птиц. У взрослого человека активный красный костный мозг составляет лишь около половины общего количества костного мозга. Другая половина приходится на долю желтого костного мозга, {79} сплошь состоящего из жировой ткани. Однако хирургическая практика показывает, что костный мозг — весьма подвижная система: желтый костный мозг может замещаться активным красным. Желтый костный мозг находится главным образом в трубчатых костях, а красный — в телах позвонков, ребрах, грудине, тазовых костях. В среднем у человека в скелете на долю костного мозга приходится около 45% веса скелета, а в отдельных костях и больше. В частности, в грудных и поясничных позвонках на долю костного мозга приходится до 70% веса позвонков.
У собаки — около 80% активного красного костного мозга и около 20% желтого. Примерно 30% красного костного мозга сосредоточено в позвоночном столбе, около 17% в ребрах и грудине, и около половины общего количества костного мозга приходится на кости передних и задних конечностей.
Подобного рода данных в отношении содержания красного и желтого костного мозга у других животных имеется очень мало. Известно, что у новорожденных млекопитающих весь костный мозг красный. А между тем точное знание соотношения красного и желтого костного мозга у различных видов животных в их онтогенезе имеет очень важное значение для оценки продуктивности костного мозга как очага кроветворения, синтеза гемоглобина.
В целом же костный мозг весьма своеобразная ткань в организме наземных позвоночных животных, которая представляет собой эмбриональную ткань и сохраняет свой эмбриональный характер в течение всей жизни особи. Может быть, необходимость сохранения нежной эмбриональной ткани и была одной из причин, обусловивших расположение ее в самых глубоких недрах организма, в плоских и трубчатых костях.
Поскольку данных о количественной характеристике костного мозга в литературе имеется мало, то судить о его продуктивности можно пока на основании определения количества крови в организме и точного учета важнейших компонентов крови — гемоглобина и эритроцитов. Точный учет количества гемоглобина и его носителей {80} — эритроцитов — важен. Во-первых, эритроциты составляют основную массу клеток в крови любого животного. Во-вторых, срок существования эритроцитов у разных представителей позвоночных различен. Поэтому, чтобы судить о продуктивности костного мозга, надо знать и темпы обновления форменных элементов крови, пребывающих в сосудистом русле.
Данные, характеризующие особенности эритроцитов крови — форму, размеры и количество в единице объема (обычно в кубическом миллиметре крови) у представителей различных групп позвоночных, представлены в таблице 3. При их сопоставлении обращает внимание определенный параллелизм изменения количества эритроцитов в единице объема и их размеров у рыб (водных животных) и у остальных позвоночных (наземных животных).
Действительно, среди разных групп рыб хрящевые обладают небольшим количеством эритроцитов — около 150 тыс. в 1 мм3 крови — крупных размеров (в среднем 1000 μ3), у высших представителей — костистых рыб — их больше — около двух млн. эритроцитов, но они более мелкие (в среднем 167 μ3). Подобная же тенденция наблюдается и у наземных позвоночных. В этом случае имеет место более широкий диапазон изменений. У хвостатых амфибий также малое количество эритроцитов; немного более 100 тыс., но они достигают гигантских размеров — 11500 μ3. У млекопитающих — высших представителей позвоночных животных — эритроцитов в 1 мм3 крови 8,5 млн., а средний объем эритроцитов равен примерно 70 μ3.
Можно только высказать предположение о причинах, обусловивших проявление этого параллелизма у рыб и остальных позвоночных. Не исключено, что повышение подвижности — одна из основных причин, обусловивших эволюцию клеток красной крови в сторону уменьшения их объема. В результате этого увеличилась поверхность эритроцитов, что является весьма существенным моментом в механизме транспорта кислорода.
Однако совершенно неясен вопрос о причинах, обусловивших появление гигантских эритроцитов в крови хвостатых амфибий, которые видимы даже простым глазом. Почему понадобились эритроциты-гиганты первым выходцам на сушу?
| {81} |
Таблица 3. Некоторые параметры эритроцитов крови различных групп позвоночных животных
Животные |
Количество эритроцитов, в 1 мм крови в млн. |
Объем одного эритроцита, в μ3 |
Количество гемоглобина в одном эритроците в % |
Концентрация гемоглобина в эритроцитах, в % |
Истинная концентрация гемоглобина в эритроцитах в % (без объема ядра) |
Кислородная емкость крови в объемных % |
Млекопитающие |
8,41 |
66,0 |
20,0 |
32,7 |
— |
19,0 |
Птицы |
2,70 |
138,0 |
40,0 |
28,8 |
34,8 |
18,5 |
Рептилии |
0,98 |
336,0 |
97,5 |
28,9 |
33,3 |
12,0 |
Амфибии бесхвостые |
0,49 |
670,0 |
179,0 |
27,5 |
30,5 |
12,0 |
Амфибии хвостатые |
0,11 |
11300,0 |
269,0 |
24,0 |
32,0 |
10,0 |
Рыбы костистые |
1,96 |
167,0 |
36,0 |
21,3 |
25,5 |
8,8 |
Рыбы хрящевые |
0,15 |
1025,0 |
209,0 |
21,9 |
25,4 |
6,5 |
Круглоротые |
0,14 |
1530,0 |
318,0 |
21,0 |
24,0 |
6,3 |
Эритроциты хвостатых амфибий почти в 10 раз крупнее эритроцитов хрящевых рыб, и примерно в 170 раз крупнее среднего эритроцита млекопитающих. Если продолжить сопоставление особенностей эритроцитов, то с момента появления круглоротых, у которых средний объем эритроцита около 1500 μ3, до высших костистых рыб объем эритроцита уменьшился лишь в 10 раз.
Несмотря на эти рельефные сопоставления, биологический смысл появления гигантских эритроцитов хвостатых амфибий остается неясным. Некоторые биологи считают хвостатых амфибий «неудачниками эволюционных экспериментов». Однако против такой оценки можно возражать. Если хвостатые амфибии существуют до наших дней, значит они вовсе уж не такие «неудачники». Более того, имеются основания считать хвостатых амфибий «удачниками», поскольку они начали успешное освоение суши, несмотря на трудности перехода в «другое» гравитационное поле.
Имеется еще одна особенность эритроцитов крови наземных позвоночных животных по сравнению с водными: все они обладают более высокой концентрацией гемоглобина по сравнению с эритроцитами рыб. Без поправки на объем, занимаемый ядром, которое не содержит гемоглобина, концентрация повышается с 21% у рыб до 24% у хвостатых амфибий и 33% у млекопитающих. {82}
Если вычесть объем, занимаемый ядром, то у всех ядерных эритроцитов крови наземных животных концентрация повышается до 32—35% по сравнению с 25%-ной концентрацией гемоглобина у рыб. В результате кислородная емкость крови позвоночных повышается с 6—8 объемных % у рыб до 19% у млекопитающих, то есть увеличивается почти в три раза.
Возможность адаптации к наземному образу жизни характеризует еще одно обстоятельство — длительность жизни эритроцитов, или длительность периода пребывания их в сосудистом русле. Немногочисленные данные подобного рода позволяют составить некоторое представление о «продуктивности» очагов синтеза гемоглобина.
Длительность жизни эритроцитов в сосудистом русле неодинакова у разных животных, но функциональная их эффективность у всех животных примерно одинакова. У гомойотермных животных каждый эритроцит в сосудистом русле совершает примерно одинаковое количество рециркуляции.
Так, например, эритроцит человека пребывает в сосудистом русле около 120 дней и за это время количество прохождений через капилляры легких, т. е. количество оксигенаций, составляет в среднем 160344. Эритроцит собаки совершает 177840 оксигенаций, крысы — 186200, кролика — 178860, кошки — 192990, курицы — 147150—186300.
Данные эти интересны потому, что они позволяют понять причины неодинаковой длительности жизни эритроцитов крови разных представителей позвоночных животных. Действительно, разными методами установлена длительность жизни эритроцитов крови разных животных: у кролика эритроциты находятся в сосудистом русле 45—60 дней, у мыши — 20—40, у морской свинки — 62, у кошки — 77, у обезьяны — 94—117, у снежного барана — 147, у гривистого барана — 170, у каракульской овцы — 130, у канадского оленя — 95, у тара — 165, у ламы гуанако — 225 дней.
Следовательно, длительность жизни эритроцитов как-то связана с уровнем интенсивности метаболизма данного животного. У мелких представителей млекопитающих, характеризующихся весьма интенсивным метаболизмом, эритроциты задерживаются в сосудистом русле {83} всего 40—60 дней, а у крупных представителей млекопитающих до 7,5 месяцев.
В этом плане интересны данные по эритроцитам гетеротермных животных, уровень метаболизма которых резко меняется в активный период и в период спячки. Так, например, у сурка в активный период жизни длительность жизни эритроцитов равна 36 дням, в период спячки 120; у золотистого — около 160. Птицы, у которых еще более высокий метаболизм (температура их тела превышает 41°С), эритроциты живут меньше, чем у многих млекопитающих: у кур она составляет всего лишь 28 дней, у утки — 39 дней.
В то же время длительность жизни эритроцитов у пойкилотермных животных, таких, как черепахи, составляет 11 месяцев, то есть превышает почти вдвое длительность жизни эритроцитов гомойотермных животных.
Если у гомойотермных животных число оксигенаций эритроцитов примерно одинаково за время пребывания в сосудистом русле животного (несмотря на различные сроки длительности жизни), то можно думать, что это правило распространяется и на пойкилотермных животных.
Таким образом, данные о длительности жизни эритроцитов некоторых представителей позвоночных животных доказывают, что наиболее высокая продуктивность костного мозга имеется у мелких млекопитающих и птиц (за 30—50 дней происходит полное обновление эритроцитов). Процесс смены эритроцитов продолжается непрерывно. В противном случае нарушение доставки кислорода приведет к катастрофическим последствиям. У каждого вида животных имеется свой уровень обеспеченности организма гемоглобином. Это обязательное условие сохранения определенного уровня энергетики.
Из общего количества эритроцитов, циркулирующих в сосудистом русле человека и определяемого в 26 триллионов эритроцитов, в течение каждого часа (если принять длительность жизни эритроцита равной 120 дням) разрушается 9 миллиардов эритроцитов. В результате деструкции эритроцитов, как известно, образуются желчные пигменты, выводимые из организма с мочой и калом. Железо и белковая часть молекулы гемоглобина попользуется в виде аминокислот организмом для {84} последующих синтезов гемоглобина в очагах кроветворения.
О продуктивности скелета, как очага синтеза гемоглобина и созревания эритроцитов, можно судить по данным, приводимым в таблице 4. В ней представлены данные по весовым особенностям скелета, костного мозга, крови, гемоглобина крови некоторых представителей позвоночных животных.
Продуктивность очагов синтеза гемоглобина у рыб не очень высокая, ибо на 1 кг веса тела в среднем приходится менее 2 г гемоглобина. Однако у первых выходцев на сушу количество гемоглобина на единицу веса тела удваивается, составляя в среднем 3,6 г. Обеспеченность организма гемоглобином у рептилий немного выше и составляет в среднем 3,8 г на 1 кг веса тела. Зато у птиц она в три раза больше, чем у рептилий и почти в семь раз больше, чем у рыб, и в среднем составляла 12,2 г гемоглобина на 1 кг веса тела. Таков уровень обеспеченности организма гемоглобином и у млекопитающих: в среднем 14,5 г гемоглобина на 1 кг веса тела. Это довольно высокие величины.
До недавнего времени разница в обеспеченности организма гемоглобином различных представителей позвоночных животных объяснялась тем, что рыбы как низшие представители позвоночных животных менее совершенны, а птицы и млекопитающие представляют собой наиболее совершенные формы животных.
Принцип единства организма и среды исключает возможность рассмотрения этого вопроса в такой плоскости. Любой организм достаточно совершенен, если он существует в данных условиях. Нет оснований называть тех или иных животных «неудачниками» эволюционных экспериментов, если они существуют до настоящего времени. С этих позиций низкий уровень обеспеченности организма рыб гемоглобином объясняется лишь тем, что для существования рыб этого количества гемоглобина вполне достаточно. Если бы возникла потребность в большем количестве гемоглобина, то, несомненно, нашлись бы возможности ее удовлетворить.
Первым выходцам на сушу — хвостатым амфибиям понадобились большие количества гемоглобина потому, что в новых условиях необходимо было тратить больше энергии для преодоления сил гравитации. Рыбам,
| {85} |
Таблица 4. Количество крови, гемоглобина, костного мозга и мышечного гемоглобина у некоторых видов позвоночных животных
Объект исследования |
Вес, кг |
Скелет, % |
Костный мозг, |
Кровь, % |
Гемоглобин, г/кг |
||
тела |
скелета |
кровь |
мышцы |
||||
Млекопитающие |
|||||||
Человек |
70,0 |
12,1 |
4,6 |
45,0 |
7,5 |
12,0 |
0,5 |
Собака |
6,6 |
10,5 |
3,2 |
30,5 |
9,0 |
12,4 |
|
Северный олень |
|||||||
взрослый |
70,5 |
13,0 |
5,1 |
47,0 |
12,5 |
17,5 |
3,5 |
новорожд. |
5,9 |
22,6 |
12,8 |
48,3 |
15,5 |
27,6 |
— |
Морская свинка |
|||||||
взрослая |
0,9 |
5,6 |
1,0 |
17,0 |
5,0 |
5,2 |
0,4 |
новорожд. |
0,1 |
11,5 |
2,8 |
24,5 |
8,5 |
11,3 |
— |
Птицы |
|||||||
Куры (белый леггорн) |
1,6 |
9,0 |
2,0 |
22,5 |
6,5 |
4,6 |
0,5 |
Цесарки |
1,5 |
10,0 |
2,2 |
22,0 |
9,5 |
9,1 |
1,3 |
Голубь |
0,3 |
10,2 |
2,4 |
23,5 |
10,2 |
9,3 |
1,9 |
Баклан больш. |
1,9 |
10,4 |
2,5 |
24,0 |
14,3 |
24,5 |
6,0 |
Рептилии |
|||||||
Ящерица |
0,016 |
12,0 |
— |
— |
5,0 |
4,2 |
|
Степной удав |
0,188 |
13,8 |
— |
— |
5,8 |
3,9 |
|
Амфибии |
|||||||
Лягушка бурая |
0,041 |
11,1 |
— |
— |
6,0 |
4,1 |
|
Рыбы |
|||||||
Сом |
0,960 |
6,5 |
Нет |
Нет |
4,0 |
1,8 |
|
Камбала |
0,350 |
— |
» |
» |
2,0 |
0,5 |
|
Севрюга |
6,0 |
7,8 |
» |
» |
2,8 |
3,4 |
|
обитающим в среде, где действие сил гравитации ослаблено, достаточно и того гемоглобина, который синтезируется в почках и селезенке.
У рептилий лишь слегка превысилась потребность в гемоглобине. Однако птицы и млекопитающие — теплокровные животные, развивающие большую активность и подвижность, т. е. обладают эффективными механизмами по преодолению сил гравитации и поэтому нуждаются в большом количестве кислорода и, следовательно, гемоглобина.
| {86} |
Преодоление сил гравитации в наземных условиях требует очень больших усилий. Поэтому, как уже говорилось, кроме скелета возник еще один очаг синтеза гемоглобина в мышцах тела. Мышцы стали очагом синтеза мышечного гемоглобина. У рыб лишь в отдельных мышечных комплексах синтезируется мышечный гемоглобин. Он обнаружен и у некоторых рептилий. Однако у птиц и млекопитающих лишь некоторые мышечные комплексы лишены гемоглобина. У высших представителей наземных позвоночных животных на долю мышечного гемоглобина (если за 100% взять общее количество гемоглобинов крови и мышц) приходится от 4,0% (человек) до 17% (северный олень) у млекопитающих и от 9,1% (куры) до 17,0% (голубь) у птиц (табл. 6 и 7).
Для рыб имеются данные лишь в отношении некоторых видов. В частности, у севрюги мышечный гемоглобин содержится в мышечных комплексах, приводящих в движение жаберные крышки и ротовую трубку.
Таким образом, приходится говорить не только о продуктивности синтеза гемоглобина скелетом, но и о продуктивности мускулатуры как очага синтеза мышечного гемоглобина. Несомненно, это является отражением тех энергетических затрат, которые производят различные представители наземных позвоночных животных в процессе преодоления сил гравитации.
Следует отметить, что приведенные в таблице 7 данные по «продуктивности» скелета и мускулатуры как очагов синтеза гемоглобина весьма фрагментарны. Они пока касаются лишь птиц и млекопитающих. Амфибии и рептилии пока не исследованы. Данные в отношении птиц и млекопитающих также не полные, так как они говорят об общем суммарном весе костного мозга по отношению к весу тела и весу скелета. Однако у взрослых животных одна часть костного мозга активная, красная, а другая — желтая, которая не имеет отношения к синтезу гемоглобина. В отношении некоторых животных дифференцированный учет произведен, но большинство исследованных животных в таком плане не изучено. {87}
Хотя данные о количестве гемоглобина, содержащегося в крови, дают определенное представление о «продуктивности» скелета позвоночных животных, необходимо как можно точнее определить количественно активную, кроветворную часть костного мозга и расположение ее в отделах скелета. Только в этом случае можно будет получить представление о кроветворной функции скелета в целом и различных его отделов. Последнее обстоятельство имеет весьма важное значение, так как преодоление сил гравитации у наземных позвоночных животных осуществляется разнообразными путями.
Рассмотрим способы передвижения у животных вообще и у наземных позвоночных животных в частности как конкретные механизмы преодоления сил гравитации.
В этом плане несомненный интерес представляет фундаментальная работа академика Л. А. Зенкевича «Очерки по эволюции двигательного аппарата животных». В этой работе Л. А. Зенкевич дает весьма обширную характеристику основных способов передвижения различных групп современных животных как позвоночных, так и беспозвоночных. Последнее обстоятельство является несомненно очень важным, ибо проблема передвижения, или локомоции, животных вообще до сих пор решалась в основном на примере изучения позвоночных животных. Известны исследования по так называемой биомеханике, когда изучались конкретные механизмы передвижения ряда позвоночных животных — человека, лошади, собаки и др. Л. А. Зенкевич справедливо подчеркивал, что движение — это «одна из наиболее первичных и основных функций живого вещества, при помощи которой растительные и животные организмы осуществляют наиболее полное заселение биосферы и ее использование в целях питания, защиты от врагов и неблагоприятных условий внешней среды, и в целях размножения»1. {88}
В самом деле, нельзя представить себе животный организм без движения. Выполнение таких основных жизненно важных функций, как дыхание, питание, воспроизведение себе подобных, без движения невозможно. Исключение составляют животные неподвижного образа жизни — группа так называемых сидячих животных (губки, некоторые кишечнополостные, черви и ракообразные). Однако сидячий образ жизни возможен лишь в случае подвижной внешней среды. Правда, и в этом случае имеются соответствующие структурные образования, постоянно находящиеся в движении,— различного рода щупальцы для схватывания добычи или органы аэрации, обеспечивающие организм кислородом. Более того, по-видимому, и все современные растения произошли от подвижных организмов, о чем можно судить хотя бы потому, что все низшие растения на ранних стадиях развития подвижны. Точно так же и у животных, ведущих сидячий образ жизни, личиночные стадии подвижны.
Подавляющее большинство современных животных способно передвигаться. Если сравнить органы движения у различных групп современных животных, то можно сказать, что основное направление эволюции животных организмов — обеспечение максимальной скорости передвижения во внешней среде, маневренности и экономичности процесса движения. Естественно, что это может быть достигнуто лишь при условии глубокой приспособленности структуры организма к определенным условиям существования, что все способы произвольного передвижения вызваны к жизни в процессе эволюции животных организмов.
Л. А. Зенкевич дает классификацию различных форм передвижения, подразделяя их на два типа: первый — в результате изменения общей формы тела и второй — при помощи специальных придатков тела при сохранении относительно постоянной формы тела.
Первый тип движения с помощью изменения формы тела в свою очередь подразделяется на три способа:
1. Гидравлическое движение с помощью образования псевдоподий, что достигается путем переливания протоплазмы (у одноклеточных организмов) и путем переливания полостных жидкостей (у роющих червей в случае многоклеточных животных) (рис. 12).
| {89} |
Способы передвижений | Механизмы передвижений |
Амебоидное ...... | Переливание протоплазмы |
Жгутиковое и ресничное | Колебательные движения жгутиков и ресничек |
Реактивное....... | Резкое выталкивание жидкости из организма |
Гидравлическое..... | Переливание полостных жидкостей |
Изгибание тела..... | Изгибание мембран, краевых плавников, всего тела |
При помощи конечностей | Параподии, плавники, ноги |
Полет......... | Крылья позвоночных, крылья насекомых |
2. Движение при помощи изгибания или всего тела или его краевых частей. В этом случае различают:
а) «волновое» движение в результате повторяющихся изгибаний его краевых частей (большинство червей, некоторые моллюски, рыбы, многие личинки насекомых и др.) (рис. 13);
б) «шагающее» движение (некоторые простейшие, свободноплавающие кишечнополостные, головоногие моллюски, и др.);
в) движение прыжками (некоторые моллюски, некоторые личинки насекомых, змеи и др.);
3. Реактивное движение (некоторые простейшие, свободноплавающие кишечнополостные, головоногие моллюски и др.) (рис. 14).
Второй тип движения при помощи специальных придатков тела подразделяется да два способа:
1. Движение при помощи ресничек, жгутиков, мембран (инфузории, жгутиковые, ксенофоры, некоторые турбеллярии, мужские половые клетки и личинки большинства беспозвоночных животных) (рис. 13).
2. Движение при помощи конечностей (гребное движение водных животных, шагание, бег, прыгание, парение, полет водных и наземных животных).
Однако из всех этих способов наибольшее распространение имеют гидравлическое, ресничное (или жгутиковое), волнообразное изгибание тела и при помощи конечностей. Простейшие обладают двумя основными способами передвижения — первый при помощи псевдоподий (или переливанием протоплазмы). По существу это гидравлический способ: образование псевдоподий на определенном участке тела обусловлено сжиманием
| {90} |
 |
 |
Рис. 12 |
Рис. 13 |
|
Рис. 12 Роющие черви 1 — Приапулюс каудатус, б — Галикриптус симиулезус Рис. 13. Различные формы движения у простейших а — жгутиковое движение бактерий, б — жгутиковое движение флагеллят, а — движение трипанозомы при помощи ундулирующей мембраны, г — ресничное движение инфузорий, д — поперечный разрез через жгутик или ресничку, в электронном микроскопе — девять периферических сократительных фибрилл и две срединные опорные, е — амебоидное движение |
|
эктоплазматической оболочки на остальной поверхности тела. Второй способ — при помощи ресничек и жгутиков. Псевдоподии свойственны корненожкам (амебы и др.), реснички и жгутики — инфузориям и жгутиковым.
Видоизменением ресничек и жгутиков являются многообразные мембраны, иногда позволяющие животному быстро двигаться. Эти движения чрезвычайно напоминают движение при помощи плавников у некоторых рыб и головоногих моллюсков. У простейших много и других способов, имитирующих почти все способы движения, свойственные многоклеточным животным.
Появление многоклеточных животных сопровождалось, естественно, значительным преобразованием форм движения. У низших представителей многоклеточных сохранились элементы двигательного аппарата простейших в виде ресничек или жгутиков и мионем. Однако ресничный аппарат сохранил свою эффективность только у мелких форм (менее 0,5 мм) и личинок многих беспозвоночных, тогда как мионемный аппарат превратился
| {91} |
 |
Рис. 14. Различные формы движения водных животных а — реактивное движение медузы, б — реактивное движение кальмара, в — изгибательиое движение червя — полихеты, г — изгибательное движение рыб |
в мышечную ткань беспозвоночных и позвоночных животных. Только одна группа кишечнополостных (ктенофоры) сохранила ресничный способ движения, несмотря на крупные размеры тела, но относительная скорость их движения очень мала (сотые доли миллиметра в секунду).
Губки — наиболее примитивные многоклеточные животные — совсем лишены эффективных способов передвижения, а кишечнополостные — в значительной степени. Утрата двигательной системы у предков губок связана, по-видимому, с увеличением размеров тела, что вызвало развитие механических средств защиты — гипертрофию скелета, переход к неподвижному существованию под его защитой. Подобные же изменения имели место и в эволюции кишечнополостных (сидячие формы с мощно развитым скелетом). У многих кишечнополостных имеется слабая подвижность, компенсируемая эффективными химическими защитными аппаратами — стрекательными клетками. В результате утраты двигательной функции у многих представителей иглокожих, усоногих раков, некоторых моллюсков развились механические средства защиты. {92}
Ряд представителей кишечнополостных обладает определенным типом мускулатуры, которой совершенно лишены губки. Правда, эта двигательная мускулатура весьма своеобразна и представляет собой эпителиально-мышечные клетки, образующие поверхностную систему кольцевых и продольных мышечных волокон. Эта система мышц не обеспечила быстрого передвижения, но явилась тем исходным материалом, из которого в процессе дальнейшего эволюционного развития возникла мускулатура различных групп червей и других групп животных. Количество мышц у кишечнополостных весьма небольшое. У медузы Суаnеа arctica, например, они составляют около 1 % веса тела.
Весь дальнейший процесс эволюции форм движения животных путем изменения формы тела был сопряжен с развитием мышечных комплексов, которые при сокращении меняют форму тела и мышечных комплексов-антагонистов, восстанавливающих прежнюю форму тела. Для сохранения формы тела появляются своеобразные эластичные стабилизаторы (кутикула нематод, межсегментные перегородки кольчатых червей, паренхима и мезенхима турбеллярий, немертин и пиявок, хорда хордовых животных).
У турбеллярий, или ресничных червей, появился характерный для червей кожномускульный мешок, и передвижение осуществляется посредством изгибания тела. Расположение двигательной мускулатуры носит диффузный характер; на ее долю приходится около 25% веса тела.
Немертины — активные хищники, двигающиеся путем изгибания тела, обладают способностью сильно сокращаться и растягиваться. У них мощно развитый эпидермис — предохранительное средство против повреждения поверхности тела.
Сохранению целостности покровов также способствуют громадные количества слизи, выделяемой поверхностью тела. Впрочем, слизь также способствует повышению скорости движения, уменьшая трение о воду.
Считается, что немертинообразные предки дали начало кольчатым червям, а кольчатые черви — членистоногим. Особенностью кольчатых червей является сегментация, уплотнение наружных покровов. Благодаря этому организм получил более надежную защиту, но вместе с {93} тем ограничилась подвижность тела. Это обстоятельство привело в свою очередь к уменьшению массы мускулатуры по сравнению с немертинами, локализованной в двух основных слоях — кольцевом и продольном. Причем продольная мускулатура резко преобладает над кольцевой, поскольку при уплотнении покровов ее функционирование становится ограниченным. В частности, у Nereis pelagica масса продольной мускулатуры колеблется от 31 до 39% массы тела, а кольцевой около 2,5%, у олигохет 20,3% продольная и 9,4% кольцевая. Пиявки как активные хищники обладают чрезвычайно мощной мускулатурой, достигающей 65% веса тела. На долю продольной мускулатуры приходится до 39% веса тела, кольцевой — до 12,5% и спинно-брюшной — до 18%.
Интересно, что группа роющих червей (эхиуриды, сипункулиды), передвигающиеся гидравлическим способом, обладает обширной полостью до 66% объема тела и сильной мускулатурой, вес которой меньше, чем у других червей (от 12 до 31% веса тела). В этом случае гидравлический способ движения оказался настолько механически эффективным, что требует относительно меньше мускулатуры.
Среди червей наиболее примитивный, исходный тип двигательной мускулатуры — типа кожномускульного мешка — имеется у турбеллярий. Диффузная мускулатура, пронизывающая тело по всем направлениям, дает ему большую подвижность в каждом участке, но загромождает внутренние части тела. Эволюция двигательной мускулатуры у филогенетически более молодых групп животных характеризуется тем, что внутренние части тела освобождаются от двигательной мускулатуры, которая концентрируется по периферии тела.
Л. А. Зенкевич отмечает, что имеется определенный параллелизм в характере расположения двигательной мускулатуры в виде сплошных пластов, идущих через все тело или посегментно, и «маневренностью» двигательных сокращений тела у различных групп беспозвоночных и позвоночных животных.
Однако освоение суши вызвало к жизни новые формы передвижения. Исключение составляют брюхоногие моллюски, полностью сохранившие тот способ передвижения, который свойствен им в водной среде. Все {94} остальные группы животных, освоившие сушу (паукообразные, трахейнодышащие, членистоногие и позвоночные), выработали новый способ передвижения с помощью ходных конечностей. Первые выходцы на сушу использовали комбинированный способ передвижения — сочетание движений конечностей и волнообразных изгибаний тела, характерных для рыб (первичнотрахейные и многоножки среди членистоногих, хвостатые амфибии и рептилии, а также водные млекопитающие среди позвоночных животных). Тело их имело сильно удлиненную форму.
У филогенетически более молодых групп тело укорочено, при движении сохраняется постоянная форма тела, движения осуществляются с помощью коленчатых или специальных придатков (паукообразные, насекомые, некоторые рептилии, птицы и млекопитающие).
Укорочение тела высших представителей позвоночных и беспозвоночных животных в наземных условиях привело к усилению мускулатуры конечностей у позвоночных и уменьшению числа конечностей у беспозвоночных.
Несмотря на то, что рычажный способ передвижения проявился почти у всех групп животных, начиная с корненожек, только у высших позвоночных и беспозвоночных он стал основным. На низших этапах развития он был недостаточно эффективен.
Следует иметь в виду, что у ряда современных животных не вся мускулатура, которая объединяется под сборным названием двигательная, всегда участвует в передвижении тела в пространстве. Такая двигательная мускулатура свойственна большинству червей, иглокожих, рыб, амфибий и рептилий. Однако у животных, обладающих несколькими способами передвижения, например плаванием и ползанием (некоторые планарии, коловратки) или хождением и полетом (насекомые, птицы), часть двигательной мускулатуры, обслуживающей один способ передвижения, может не принимать участия в другом и топографически обособлена (мускулатура ног и крыльев у птиц и насекомых).
Таким образом, все многообразие способов передвижения позвоночных животных изучалось исследователями и достаточно подробно систематизировано. Какой из описанных способов передвижения самый совершенный? {95}
Известно, что животное передвигается, чтобы добывать пищу, спастись от врага, уйти от неблагоприятных условий, найти представителей противоположного пола. Совершенно очевидно, что, чем быстрее скорость передвижения, тем лучше будет выполнена задача. Следовательно, в конечном счете основная цель любого из способов передвижения — сохранение благополучия особи и, тем самым благополучия вида.
Часто говорят о животных более и менее совершенных, более и менее быстроходных. Называют рекордсменов бега, полета и их антиподов — черепаху и улитку как символ медлительности. В этом утверждении есть доля правды. Если попытаться сравнить скорости передвижения в пространстве представителей различных групп современных животных, то окажется, что они весьма различны как у наземных, так и у водных животных. Ниже приведены данные, характеризующие абсолютные максимальные скорости передвижения некоторых представителей наземных и водных животных:
км/час | км/час | |||
Гепард — дикая кошка . . . | 110 | Кашалот........ | 22 | |
Антилопа сайгак..... | 70 | Тюлень........ | 20 | |
Лошадь......... | 55 | Меч-рыба ....... | 130 | |
Северный олень ..... | 40 | Тунец......... | 82 | |
Стриж.......... | до 150 | Летучая рыба ..... | 65 | |
Саранча......... | до 90 | Голубая акула..... | 40 | |
Кит-сейвал........ | 55 | Кальмар........ | 41 | |
Дельфин-белобочка .... | 44 | Осьминог........ | 15 |
Самые высокие скорости у птиц (ласточка до 150 км/час) и рыб (меч-рыба до 130 км/час). На третьем месте самое быстроходное наземное животное — гепард (110 км/час). Если взять среднюю максимальную скорость для всех млекопитающих, то она не превысит 60 км/час, тогда как у костистых рыб составит 100 км в час; примерно такая же скорость у птиц. Понимая всю условность подобной статистической обработки приведенных цифр, можно все же сказать, что рекордсменами высоких скоростей являются птицы и рыбы. {96}
Следует учесть, что наземные млекопитающие животные, в том числе и гепард, не способны развивать самую высокую скорость в течение длительного времени. По наблюдениям французского зоолога Франсуа Бульера, гепард пробежал расстояние в 650 м за 20 секунд. Средняя скорость оказалась равной 110 км/час. Гепард уже в первые секунды набирает скорость до 75 км в час. Такое ускорение недоступно пока ни одному гоночному автомобилю. Тем не менее, столь высокую скорость гепард может развивать лишь в течение очень краткого времени, измеряемого минутами. Птицы и рыбы обладают способностью сохранять высокие скорости передвижения длительное время. Известно, что стрижи в течение многих часов летают без отдыха. Многие виды рыб способны развивать высокие скорости на больших расстояниях. Следовательно, ответ на вопрос — какой из этих способов передвижения самый совершенный? — напрашивается сам собой. Самый совершенный — полет и плавание.
То обстоятельство, что птицы летают быстрее млекопитающих, бегающих по земле, еще можно понять. И те и другие относятся к категории высших представителей позвоночных животных, обладают постоянной высокой температурой тела и, следовательно, интенсивным обменом. У птиц даже температура тела более высокая (41—44°С), чем у млекопитающих (37—38°С). Но чем объяснить быстроходность рыб? Ведь рыбы — представители низших позвоночных животных — и, как утверждают некоторые биологи, являются менее совершенными организмами по сравнению с птицами и млекопитающими. Тем не менее они превосходят по скорости движения самых быстроходных млекопитающих — высших представителей класса позвоночных животных? В чем же совершенство высших позвоночных животных, если их перегоняют такие несовершенные существа, как рыбы?
Ответа на этот вопрос биологи до сих пор не дали. Они добросовестно описывают различные способы передвижения различных представителей современных животных, систематизируют их и пока этим ограничиваются. Нам представляется, что основной причиной различий в способности развивать высокие скорости у наземных животных, с одной стороны, и у водных и воздушных животных — с другой, являются силы гравитации. Они {97} проявляют свое действие в полной мере в наземных условиях и в сильной степени ослабляются (в смысле биологического эффекта) в водных и в воздушных условиях, особенно при парящем полете.
Именно с точки зрения действия сил гравитации следует рассматривать особенности структуры органов передвижения у всех основных групп современных животных и их эффективность. Ведь в конечном счете все изменения структуры органов передвижения у разных представителей позвоночных и беспозвоночных животных определяются необходимостью преодоления гравитационного поля Земли, в котором жизнь возникла и развивалась. Силы гравитации определяют характер изменения органов передвижения и энергетику организма.
Анализ особенностей передвижения животных в водной, наземной и воздушной среде показывает, что, несмотря на некоторый параллелизм в изменении структуры органов движения организма животных, имеются существенные различия в уровне энергетики организма в зависимости от особенностей среды обитания.
В главе II при рассмотрении особенностей органов движения наземных животных отмечалось, что ползание — основная форма передвижения хвостатых амфибий — наименее эффективная форма.
Бесхвостые амфибии овладели новой формой передвижения — прыжком,— несомненно более эффективным способом преодоления сил гравитации. Ранее отмечалось, какая глубокая перестройка организма потребовалась для того, чтобы овладеть этой новой формой передвижения. Для осуществления прыжков у бесхвостых амфибий должно было измениться соотношение частей скелета и мускулатуры: почти половина веса скелета и около 65% всей мускулатуры приходится на долю заднего пояса конечностей. Чтобы судить о масштабах этой перестройки организма, следует напомнить, что у амблистом (представителей хвостатых амфибий) на долю осевого скелета приходится 71% общего веса скелета, 13% — на долю переднего и 16% — на долю заднего пояса конечностей. Примерно такие же соответствия {98} имеют место у бегающих рептилий, в частности у туркестанской агамы на долю осевого скелета приходится 66% общего веса скелета, на периферический скелет — 34%. Мы не знаем, какие соотношения веса различных отделов скелета и мускулатуры имели место у летающих рептилий. Следует подчеркнуть, что рептилии среди наземных позвоночных были первыми представителями, которые успешно преодолели силы гравитации и смогли подняться в воздух на длительное время.
Успешно преодолели силы гравитации большинство птиц и летающие млекопитающие. Полет у тех и других осуществился в результате глубокой перестройки организма подобно бесхвостым амфибиям. Не будет преувеличением сказать, что вся структура организма птиц подчинена идее полета. Почти половина веса их скелета и свыше 70% общего веса мускулатуры у крачки обыкновенной приходится на долю переднего пояса конечностей. Аналогичная картина наблюдается у летающих млекопитающих: у малого бухарского подковоноса 53% веса скелета и 71% веса мускулатуры приходится на долю переднего пояса конечностей. Это высшие величины относительного веса скелета и мускулатуры переднего пояса конечностей. Впрочем, не исключено, что у колибри эти величины могут быть превышены. В связи с особенностями полета у этой птицы самое крупное сердце, относительный вес которого составляет почти 3% от веса тела! Ни одно из современных животных не обладает таким крупным сердцем. Однако у плохо летающих птиц (куры, цесарки) относительный вес скелета переднего пояса снижается до 17,5 и 20,1% и вес мускулатуры до 43 и 45%. Зато на долю заднего пояса конечностей приходится 39 и 46% веса скелета и 39 и 38% мускулатуры. Следует учесть, что речь идет в основном о бегающих птицах, в жизни которых полет играет ограниченную роль.
У безкилевых птиц (страусы, киви) эти величины должны быть еще более резко выражены, ибо у них полет как форма передвижения отсутствует. Представляют интерес пингвины, чрезвычайно приспособившиеся к водному образу жизни. Хотя они не летают, но при нырянии используют крылья как весла. У них на долю мускулатуры приходится 41% и на скелет переднего Пояса конечностей — 25%. {99}
Приведенные данные об особенностях костно-мышечной системы, осуществляющей подвижность животного, свидетельствуют о том, что наиболее эффективные формы передвижения, а следовательно, и наиболее эффективные пути преодоления сил гравитации предполагают исключительно глубокие изменения структуры организма вообще и костно-мышечной системы в особенности. Это значит, что силы гравитации в условиях наземного существования определяют пути эволюции наземных животных, становясь одним из самых мощных факторов эволюции.
Первые выходцы на сушу среди позвоночных животных — амфибии — столкнулись с действием резко возросших сил гравитации. Рептилии, птицы и млекопитающие в полной мере приспособились к этому фактору, освоив не только сушу, но и воздушное пространство, чего не могли сделать амфибии.
В связи с этим, может быть, следует отметить, что до сих пор биологи рассматривают переход от водного к наземному образу жизни как рядовой этап, не представляющий чего-либо принципиально нового, лишь как простую смену механизмов передвижения. И несмотря на то, что при этом иногда подчеркивается, что на суше животные оказались под жесточайшим контролем «возросшего» веса тела, основной смысл адаптации к наземным условиям усматривался лишь в выработке устойчивости движения. Это, конечно, очень важное обстоятельство, однако только с этих позиций нельзя понять многих явлений из жизни наземных позвоночных. В. Б. Суханов прогресс в адаптации амфибий излагает следующим образом:
«Первые наземные позвоночные — стегоцефалы (мастодонозавр) передвигались очень медленным шагом. Конечно, это мало их устраивало: скорость — важнейший фактор в отнюдь не мирном сосуществовании животных. Но вопрос стал так: хочешь прибавить скорости — позаботься о том, чтобы сам характер движения менялся в сторону рыси, иначе появятся моменты, когда придется балансировать на двух лапах одной стороны, а это не шутка — попробуйте представить себе в таком положении, например, черепаху. Чем больше скорость, тем более и более в унисон должны работать диагонально расположенные лапы. {100}
Рысь появилась только у пресмыкающихся, но и среди земноводных не все двигаются шагом, хотя бы и рысеобразным. Червяки, лишенные ног в связи с роющим образом жизни, стали ползать на манер дождевых червей. Вернувшись в воду, тритоны плавают по-рыбьему.
Иначе все пошло у лягушек. Предки их, по каким-то причинам потерявшие хвост, тоже вернулись в воду, но так как к тому же и тело у них было укорочено, то плавать, как рыбы, они не смогли и были вынуждены отталкиваться одновременно обеими задними лапами. Приспособление оказалось удачным и позволило даже на суше передвигаться прыжками. Некоторые бесхвостые амфибии забирались на деревья (квакша), а одна из них даже отважилась на прыжки с «парашютом» (веслоногая лягушка)»1.
В. Б. Суханов отмечает, что «приспособление оказалось удачным», и поэтому животное смогло даже на суше передвигаться прыжками. Дело в том, что амфибии никогда окончательно не покидали воду, так как угроза гибели от высыхания удерживала их в прибрежной зоне водоемов. Но если почти все ресурсы организма служат для реализации прыжка, то как же можно говорить, что это приспособление позволило передвигаться «даже на суше»? Предки лягушек не «по каким-то причинам» потеряли хвост, а в связи с необходимостью более эффективного преодоления сил гравитации.
О том, что именно необходимость преодоления сил гравитации была основной причиной потери хвоста предками лягушек, говорят и другие факты. Анализ особенностей передвижения животных в водной, наземной и воздушной среде показывает, что наиболее значительные траты энергии производятся животными в наземных условиях. Известно также, что у хороших скороходов, бегунов и прыгунов прекрасно развиты органы аэрации и сердечно-сосудистая система, кровь богато оснащена гемоглобином, т. е. имеются все необходимые предпосылки для эффектного обеспечения кислородом организма, совершающего большую работу. В то же время у рыб, способных развивать не меньшие скорости при передвижении, чем наземные позвоночные, оснащенность организма гемоглобином значительно меньше, а соответственно {101} и менее интенсивна деятельность органов аэрации и гемодинамики. Следовательно, рыбы преодолевают те же трудности, что и их наземные собратья, но только более экономным путем!
Птицы в этом отношении занимают особое положение. Их пребывание в воздушной среде все же ограничено, даже в том случае, если они добывают там пищу (стрижи, ласточки) или поглощают нектар цветов (колибри), так как на ночь и на время сооружения гнезда они приземляются. Именно потому, что им практически приходится обитать в двух средах, они обладают более сложными формами адаптации, чем рыбы, которые обитают в водной среде и адаптированы только к ней. Поэтому и формы адаптации у них выражены более четко по сравнению с истинно наземными животными.
По-видимому, сложными формами адаптации можно объяснить те особенности в энергетике организма, которые наблюдаются у различных представителей животных, если сравнивать интенсивность обмена веществ не только в покое, но и в момент максимальной деятельности. Известно, что одна из особенностей сравнительных исследований по энергетике организма — утверждение, что сравнивать уровень энергетики организма у разных животных можно, измеряя лишь обмен покоя или базальный метаболизм. Но в ряде случаев обмен покоя — состояние организма, весьма трудно достижимое, а в некоторых случаях и просто недостижимое. Действительно, практически не удается получить обмен покоя у землеройки — самого мелкого и вместе с тем наиболее активного животного из млекопитающих — или у колибри — также самой мелкой и, по-видимому, наиболее активной из современных птиц.
Именно поэтому было выдвинуто положение о необходимости изучения не только обмена покоя, но и максимального обмена веществ, имеющего место при самой интенсивной деятельности. Подобных данных очень мало, однако анализ их показывает, что рыбы способны усиливать обмен веществ лишь в 2—4 раза и очень редко {102} в 6—7 раз (проходные рыбы, главным образом, лососевые рыбы), млекопитающие в 7—12 раз, а птицы в 25—50 раз.
Нам представляется, что эти различия обусловлены тем, что рыбы как обитатели водной среды, где биологический эффект действия сил гравитации резко ослаблен, затрачивают наименьшее количество энергии для преодоления их действия. Значительно больше энергии для преодоления сил гравитации затрачивают млекопитающие, обитающие в наземной среде, где действие этих сил проявляется в полной мере. Максимальное количество энергии затрачивают птицы, обитающие в двух средах — наземной и воздушной. Впрочем, непревзойденными рекордсменами являются летающие насекомые, которые обладают способностью увеличивать интенсивность обмена веществ в полете в 100 раз и более по сравнению с обменом покоя.
Следует подчеркнуть, что всем организмам приходится затрачивать огромные усилия для того, чтобы преодолеть действие сил гравитации и оторваться от поверхности земли. Но не все представители животных, хорошо освоивших сушу (рептилии, птицы, млекопитающие и насекомые) успешно это осуществляют. Лишь некоторые из них эффективно преодолели действие сил гравитации и длительное время могли находиться в воздухе. Это — летающие ящеры, ранее вымершие, подавляющее большинство современных птиц и летающие млекопитающие. Остальные могут лишь на момент отрываться от поверхности земли во время прыжков. В этой категории животных есть свои рекордсмены, обладающие способностью отрываться на максимально большую высоту.
Несомненно, рекордсменами по прыжкам в высоту опять-таки являются насекомые. Если же говорить о позвоночных животных, то среди этой группы животных, по-видимому, первое место принадлежит дельфинам, способным прыгать в высоту до 6 м, некоторым представителям проходных рыб (например, лосось прыгает вверх до 3,5 м) и кенгуру — до 3 м. Рекордсмен мира по прыжкам в высоту Валерий Брумель поставил мировой рекорд в 1962 г., прыгнув на высоту 2 м 28 см. Однако, если бы Брумель обладал такой же мощью, как насекомые, то мог бы прыгнуть на высоту до 100 м. Но человек слабее насекомых. У насекомых наиболее эффективное {103} дыхание по сравнению со всеми современными животными. Трахейная система отличается от органов дыхания, свойственных другим группам животных, тем, что доставляет кислород непосредственно клеткам и тканям тела. У других групп животных пути поступления кислорода в недра организма более сложные, вследствие чего нарушение в любом из звеньев системы снабжения может привести к кислородному голоданию. Насекомые не знают, что такое кислородное голодание — они всегда получают его в изобилии.
Вопрос о передвижении животных до сих пор сводится к изучению конкретных механизмов передвижения. Делались попытки классифицировать различные формы передвижения — бег, полет, прыжки, ползание, плавание — и характеризовать конкретные механизмы передвижения — конечности как системы рычагов для передвижения, крылья как несущие поверхности и т. д. Однако на все это многообразие механизмов перемещения животных нужно смотреть не с точки зрения простой классификации или систематизации механизмов перемещения животных в пространстве, что само по себе важно и нужно, а с точки зрения эффективности преодоления животными сил земного притяжения. Тогда вопрос о том, как передвигаются животные, будет сведен к равноценному вопросу: как они преодолевают силу земного притяжения, или силы гравитации? Тогда все многообразие способов передвижения животных можно будет рассматривать с одной, на наш взгляд единственно правильной, точки зрения. Ведь практически передвижение сводится именно к преодолению сил гравитации. У одних это происходит весьма эффективно, у других менее эффективно. Однако во всех случаях преодолеваются силы гравитации. И если у тех или иных животных механизмы преодоления сил гравитации менее совершенны, чем у других, то обладателей этих механизмов они вполне удовлетворяют, ибо эти животные существуют так же, как и обладатели более совершенных механизмов.
Можно также считать, что без сил гравитации не было бы и такого разнообразия форм передвижения животных, которое мы наблюдаем у современных животных.
| {104} |
Глава IV
Принцип единства организма и среды как один из самых фундаментальных законов биологии предполагает наличие многообразных форм приспособления, возникающих в процессе эволюции жизни на нашей планете. Исходя из этих позиций, следует анализировать специфические формы адаптации животных, освоивших сушу, при вторичном возвращении в водную среду. Поэтому особенно важная задача — изучение конкретных форм адаптации к водным условиям существования животных, ранее приспособившихся к жизни на суше, которые обеспечили в водной среде нормальное течение основных жизненно важных функций — дыхания, питания, воспроизведения себе подобных.
В водных условиях, по-видимому, наибольшие трудности связаны с адаптацией функции дыхания и воспроизведения себе подобных. Мыслимы два пути адаптации к водной среде — переход на использование растворенного в воде кислорода, как это происходит у рыб (и у предков наземных животных), или использование кислорода атмосферы. Однако, поскольку имеет силу принцип необратимости эволюции, впервые сформулированный в 1893 г. бельгийским палеонтологом Доллб, в данном случае речь может идти не о восстановлении жаберного типа дыхания, а об адаптации наземной формы дыхания. В этом Случае необходимы периодические контакты с атмосферой. Ритм определяется мощностью резервов кислорода, которые имеются в организме.
В природе известны формы адаптации к водной среде, позволяющие использовать для дыхания растворенным в воде кислород. Так дышут все первичноводные организмы, личинки многих насекомых и амфибий. На самых {105} ранних этапах эмбрионального развития дышут растворенным кислородом эмбрионы рептилий, птиц и млекопитающих. Однако все вторичноводные позвоночные сохранили воздушный тип дыхания, то есть используют для дыхания кислород атмосферы. Природа избрала наиболее рациональный в данной ситуации путь. Каковы же конкретные формы адаптации организма вторичноводных животных, которыми обладают эти животные?
Совершенно очевидно, что вторичноводные животные—это не однородная группа. Для одних характерно временное пребывание в водной стихии, связанное лишь с добыванием пищи (нутрия, ондатра и др.), для других—возвращение на сушу на период размножения (ластоногие), для третьих—окончательный разрыв с сушей и сохранение лишь периодических контактов с атмосферой (китообразные). Следовательно, среди вторичноводных животных не может быть единообразия в формах адаптации к водной среде обитания, и нет ни одного вида животных, который полностью порвал бы не только с сушей, как это сделали китообразные, но и с воздухом. Воздух оказался той единственной нитью, которая связывает китообразных с их прежней средой обитания. Все формы адаптации в водной среде были направлены на то, чтобы эта единственная нить не оборвалась.
Естественно, что адаптация касалась и других структур организма, в частности, формы тела, способов передвижения, действия рецепторов, адаптированных для функционирования в наземных условиях, и т. д. Однако эти формы адаптации изменялись лишь в той мере, в какой это необходимо было для сохранения возможности нормального выполнения основных жизненно важных функций организма — дыхания, питания и размножения.
Целесообразно будет начать рассмотрение форм адаптации вторичноводных животных к водной среде обитания с группы, наиболее приспособленной к жизни в воде, то есть с китообразных. Описание китообразных обычно сводится к констатации хорошо обтекаемой рыбообразной формы тела, превращения передних конечностей в ласты, редукции задних конечностей до рудиментарных {106} косточек. Что же касается конкретных механизмов адаптации, относящихся к реализации функции дыхания китообразных, способности выключать внешнее дыхание на сроки разной длительности, то эти вопросы почти не разработаны:
Длительность | |
Птицы | |
Кайра.............. | 6—12 |
Пингвин............. | 7 |
Млекопитающие | |
Ондатра............. | 12 |
Бобр .............. | 15 |
Серый тюлень........... | 15 |
Сельдяной полосатик ....... | 30 |
Кашалот............. | 60—74 |
Бутылконос ........... | 120 |
Человек | |
Непрофессионал ......... | 1 |
Профессионал (ловец жемчуга) ... | 2,5 |
До сих пор нет ясного представления о сроках выключения внешнего дыхания различных представителей китообразных, особенно крупных. В фундаментальных сводках приводятся данные, согласно которым кашалот и бутылконос выключают внешнее дыхание на полтора часа, сельдяной кит-полосатик до 30 минут.
В то же время нет никаких физиолого-биохимических данных, которые бы подтверждали справедливость этих утверждений. Оправданием неопределенности в этом вопросе являются огромные размеры животных, которые при современной технике исследований почти недоступны для изучения. Это обстоятельство, конечно, сильно затрудняет выявление конкретных форм физиолого-биохимических адаптации крупных китообразных. Мелкие китообразные представляют неоспоримые преимущества. Данные, полученные при изучении морфологических и физиолого-биохимических особенностей мелких китообразных, наиболее достоверны.
Установлено, что длительность ныряния дельфинов — представителей мелких китообразных — весьма ограничена. Если дельфин находится на суше, то он может сутками {107} лежать и дышать, во всяком случае с ним не происходит той быстрой трагической развязки, которая обязательно сопровождает пребывание на суше крупных китообразных, погибающих от собственной тяжести. В газетах и журналах разных стран часто появляются сообщения о гибели китов, оказавшихся на суше в силу тех или иных обстоятельств. В качестве иллюстрации можно привести заметку, опубликованную в одном из номеров журнала «За рубежом» в 1968 г., в которой сообщалось о необычной трагедии, разыгравшейся на Атлантическом побережье Франции. 15 кашалотов, несмотря на отлив, оказались на мелководье. Вода убывала с такой быстротой, что увлекшиеся охотой киты не сумели вернуться на глубокое место. Рыбаки из Жоди (недалеко от Бреста) обнаружили их беспомощно лежащими в прибрежном иле. Попытки спасти кашалотов оказались безрезультатными; слишком велик, вес животных, чтобы их можно было отправить в океан. Кашалоты погибли до того, как начался новый прилив.
Дельфин, например белобочка, если ему закрыть дыхало, может спокойно лежать не более двух минут. По прошествии двух минут или даже раньше начинается бурная реакция. Такая же картина наблюдается в случае другого вида черноморского дельфина — афалины. Афалина с закрытым дыхалом лежит спокойно не более четырех минут. Возможно, что в воде, этот срок может удвоиться или даже утроиться. Однако и в этом случае длительность выключения дыхания будет у белобочки 6 минут, у афалины — 12 минут. Эти величины далеки от 30 минут и, тем более, от 1—2 часов. Именно поэтому вызывают сомнения сроки длительности ныряния крупных китообразных, приводимые в некоторых сводках, и к ним следует относиться критически.
Имеется и еще одно обстоятельство, вынуждающее брать под сомнение такие длительные сроки ныряния, как 30 минут и полтора — два часа. Известно, что организм высших позвоночных животных и человека обладает весьма малыми резервами кислорода. На всех этапах эволюции животного мира благополучие организма {108} обеспечивалось созданием резервов важнейших компонентов, таких как жиры, углеводы, соли, витамины и другие вещества, необходимые для поддержания нормального уровня обмена. Некоторые из этих резервных веществ накапливаются в больших количествах. У некоторых видов животных жиры составляют от 30 до 50% веса тела животного, как это имеет место у гетеротермных животных (сурок) и водных млекопитающих (тюлень). Однако кислород — важнейший элемент, абсолютно необходимый для нормальной жизнедеятельности, без которого наземные организмы могут обходиться одну-две минуты — не имеет сколько-нибудь значительных резервов в организме.
Природа не создала значительных резервов кислорода в организме на протяжении почти всей истории развития позвоночных животных. По-видимому, имелись фундаментальные причины, которые на всех этапах филогенеза позвоночных животных мешали созданию резервов кислорода в организме. Только крупные китообразные, если верить визуальным наблюдениям, имеют резервы кислорода, позволяющие им нырять на сроки от 30 минут до двух часов.
Но эти сроки мало вероятны и по другой причине. Для организма имеет жизненно важное значение не только химический состав воздуха, но и его физическое состояние. Впервые это было показано А. Л. Чижевским в начале 20-х годов нашего века.
Одна из важнейших особенностей воздуха — определенная насыщенность его электрическими зарядами, вследствие чего молекулы кислорода переходят в ионизированное состояние. А. Л. Чижевский показал, что если в воздухе нет ионизированного кислорода, то все подопытные животные довольно быстро погибают. Эти исследования, которые до сих пор не учитывают физиологи и биохимики, проливают свет на причины, обусловившие отсутствие в организме животных значительных резервов кислорода.
Дело в том, что ионизированный кислород существует очень непродолжительное время — около десяти минут, после чего заряд теряется, и кислород превращается в неактивную форму. Именно по этой причине организм животного лишен возможности иметь сколько-нибудь существенные запасы ионизированного кислорода {109} отрицательной полярности1. Молекулы кислорода, не имеющие отрицательного электрического заряда, как показали опыты А. Л, Чижевского, не в состоянии поддерживать нормальную жизнедеятельность организма животных.
Отрицательно заряженный ион кислорода, обладая девятью электронами, напоминает по своим свойствам фтор. Кроме того при ионизации кислорода изменяется спин его ядра. По-видимому, эти тончайшие физические явления, происходящие на электронном и ядерном уровне, играют решающую роль в эволюции животного мира, в результате чего даже у высших представителей позвоночных животных нет значительных резервов кислорода. Ведь проблема получения кислорода и его резервирования не могла представить в процессе эволюции животных непреодолимых трудностей. Однако накопление мало активного кислорода, видимо, не решало проблемы создания резервов кислорода. Вследствие этого адаптивные изменения пошли по линии резкой интенсификации деятельности органов аэрации и гемодинамики, что и нашло наиболее совершенное выражение у высших представителей позвоночных животных — птиц и млекопитающих.
Все, что известно о резервах кислорода, получено главным образом в опытах на мелких китообразных и ластоногих. Особенность дыхания водных млекопитающих заключается в том, что у них весьма понижена чувствительность дыхательного центра к накоплению угольной кислоты в крови. У наземных млекопитающих даже незначительные изменения в содержании угольной кислоты в крови оказывают влияние на частоту дыхания. Увеличение содержания углекислоты в крови усиливает вентиляцию легких.
Кроме того, для всех водных млекопитающих характерно еще одно явление, касающееся деятельности сердечно-сосудистой системы — так называемая брадикардия, или урежение частоты сокращений сердца, о чем судят по изменению частоты пульса. Брадикардия свойственна и таким неводным млекопитающим, как человек. Правда у прекрасных ныряльщиков — тюленя, пингвина, {110} бобра — урежение пульса достигает 8—13-кратных величин, а у дельфина и человека лишь 2,5.
Поскольку явление брадикардии свойственно всем млекопитающим и птицам, биологический смысл его, по-видимому, состоит не только в более экономном расходовании кислорода в период ныряния, но также и в уменьшении энергетических трат, необходимых для пребывания во взвешенном состоянии. Для дельфина характерна высокая кислородная емкость крови, а соответственно и высокая концентрация гемоглобина. Она достигает высшей величины, известной для современных животных — 31—32 объемных процента кислорода, т. е. свыше 23 г—% гемоглобина. Но количество крови в организме дельфина-белобочки оказалось средней величиной. Определение объема крови у живых дельфинов с помощью витальной краски трипан-бляу и методом перфузии показало, что количество крови в организме дельфина-белобочки равно примерно 8% веса тела. Это значительно меньше, чем предполагалось ранее. В частности Шоландер, рассчитывая вероятные резервы кислорода в организме дельфина, исходил из предположения, что общее количество крови в организме дельфина составляет около 15% веса тела.
Количество крови в организме дельфина оказалось равно количеству крови в организме домашних овец и других сельскохозяйственных животных. Это обескураживало. Данные были не вполне понятными, ибо a priori казалось, что выключение внешнего дыхания даже на несколько минут (а дельфин-белобочка ныряет на 2—4 минуты) должно быть обеспечено не только высокой концентрацией гемоглобина крови, но и большим количеством крови.
Еще ранее было высказано предположение, что у китообразных должно быть много гемоглобина, много крови и должны быть мощно развитые органы кроветворения. Подтверждением справедливости этих предположений был вес скелета крупных китообразных, достигающий 20—30% веса тела. Относительный вес скелета наземных млекопитающих, например сельскохозяйственных животных, составляет 9—13% веса тела. Считалось, что эти данные подтверждают справедливость предположения о наличии большого количества крови в организме китообразных и мощного развития органов кроветворения.
| {111} |
Известно, что органом кроветворения у наземных позвоночных животных является активная кроветворная часть скелета — костный мозг. Логично было ожидать в мощно развитом скелете крупных китообразных много костного мозга. У наземных млекопитающих количество костного мозга составляет до 45% общего веса скелета, т. е. почти половина веса скелета приходится на долю костного мозга. Мы полагали, что и у дельфинов вес скелета будет составлять, по крайней мере, 15—20% веса тела. С целью проверки справедливости этого предположения и было проведено определение количества крови в организме черноморского дельфина-белобочки наряду с определением веса скелета, костного мозга и мускулатуры. Вместо ожидаемого большого количества крови в организме дельфина было обнаружено, как указывалось выше, весьма умеренное количество крови и еще более легкий скелет, вес которого составлял всего лишь 7—9% общего веса тела дельфина. Последующие опыты показали, что при более тщательной очистке скелета дельфина-белобочки и дельфина-азовки относительный вес скелета равен всего лишь 5—6% веса тела дельфина, а количество костного мозга составляет не более 2% веса тела.
В точности этих данных нет оснований сомневаться. Тогда возникает вопрос — а как относиться к литературным данным о весе скелета крупных китообразных, о которых речь шла выше? Возможно, большой вес скелета крупных китообразных является своеобразной формой адаптации к размерам тела, обеспечивающей прочность конструкции организма китов. А может быть, скелет оказался таким тяжелым вследствие недостаточно тщательной очистки? Вопрос этот пока остается открытым.
Однако анализ литературных данных, касающихся веса скелета, показывает, что, по-видимому, ближе к истине второе допущение. Почти во всех работах говорится, что на ребрах оставались куски мяса. А ведь, кроме кусков мяса, есть сухожилия, связки, хрящи, спинной мозг, которые конечно не учитывались, поскольку их трудно удалять. Ведь промысловики интересовались весом различных частей тела кита с технологической точки зрения выхода ценных продуктов — жира и мяса. Для {112} таких целей грубый способ очистки скелета вполне подходит. Однако для решения теоретических проблем он явно непригоден.
Полученные нами данные о весе скелета и костного мозга китообразных свидетельствуют о тем, что переход к водному образу жизни был связан с определенной редукцией скелета, выразившейся не только в потере трубчатых костей конечностей, но и других частей скелета, что нашло свое выражение в снижении относительного веса скелета до 5% веса тела.
Как объяснить это явление? В наземных условиях чем активнее животное, тем тяжелее его скелет, тем больше костного мозга и гемоглобина. В частности, у северного оленя вес скелета без рогов и копыт составляет около 13% веса тела, у горного барана архара около 16%, у морской свинки около 5%. То есть имеется почти пропорциональная зависимость между активностью животного и весом скелета: морская свинка ведет почтя неподвижный образ жизни и имеет очень легкий скелет; прекрасные скороходы северный олень и дикий баран архар, развивающий скорость до 60 км в час, имеют скелет очень тяжелый.
Но дельфин также является прекрасным пловцом и с успехом состязается в скорости с пассажирскими пароходами. Следовательно, он тоже должен иметь тяжелый скелет! В действительности же вес его скелета того же порядка, что и у малоподвижной морской свинки, ведущей наземный образ жизни. Может быть только одно объяснение этим фактам: все изменения определяются сменой среды обитания, переходом предков дельфина к водному образу жизни, то есть в «гипогравитационную» среду, где действие сил гравитации резко ослаблено. Скелет как основная структура, предназначенная для преодоления сил гравитации в наземных условиях, при переходе к водному образу жизни стал испытывать меньшую нагрузку. Это и привело к его значительной редукции.
Однако с первого взгляда такие изменения в скелете при переходе к водному образу жизни кажутся не вполне рациональными. Ведь скелет — это не только каркас, определяющий форму организма, не только система рычагов для передвижения животного, но и самый мощный очаг кроветворения, синтеза гемоглобина. А {113} гемоглобин нужен в водных условиях может быть в большей степени, чем в наземных, ибо жизнь в воде связана с необходимостью выключения внешнего дыхания на период ныряния.
Следовательно, на период ныряния необходимо создавать резервы кислорода, что возможно только при богатом оснащении организма гемоглобином! Природа разрешила это противоречие совершенно оригинальным способом: был создан новый мощный очаг синтеза гемоглобина в виде поперечно-полосатой мускулатуры. Такой путь разрешения противоречий не представляет собой чего-либо необычного. Известно, что любая клетка организма способна синтезировать гемоглобин, если в этом будет необходимость. В частности, известны гемоглобины нервной ткани, жировой ткани, даже гемоглобины зеленых растений (в корневых клубеньках бобовых растений имеется гемоглобин).
Таким образом, вопрос сводится к выявлению причин ограничения масштаба деятельности скелета как очага синтеза гемоглобина и стимулирования биосинтеза гемоглобина в мышцах тела. По-видимому, приведенное выше объяснение правильно: в водных условиях нагрузка на скелет снизилась и автоматически уменьшились масштабы синтеза гемоглобина в скелете. Однако потребность в кислороде не снизилась! Мышцы тела и в воде совершают большую работу, поэтому и потребность в кислороде остается высокой. Для этой цели в мышечных клетках синтезируется особый вид гемоглобина, обладающий более высоким сродством к кислороду, чем гемоглобин крови, который и выполняет задачу обеспечения мышц кислородом.
Мышечный гемоглобин гладких и поперечно-полосатых мышц имеется у многих представителей не только позвоночных, но и беспозвоночных животных. Однако различие состоит в том, что китообразные развили в себе способность синтезировать мышечный гемоглобин в концентрациях, во много раз превышающих уровень, свойственный наземным млекопитающим. Действительно, {114} определение мышечного гемоглобина в мышцах осевого скелета дельфина показало, что его концентрация достигает 3500 мг—%, то есть примерно в 10 раз больше, чем у наземных млекопитающих.
Одна из отличительных особенностей мышечного гемоглобина — неодинаковая концентрация его в разных мышечных комплексах одного и того же животного. Уровень концентрации мышечного гемоглобина определяется степенью функциональной нагрузки на тот или иной мышечный комплекс. Это подтверждают приводимые ниже данные концентрации гемоглобина в мг—% в различных мышцах млекопитающих и птиц:
Сердце |
Диафрагма |
Мышцы |
Груд- |
Мышцы |
Мышцы |
|
Дельфин-белобочка |
1240 |
2800 |
— |
— |
2100 |
3600 |
Тюлень ..... |
— |
— |
— |
— |
— |
5000—10000 |
Ондатра...... |
1550 |
1790 |
— |
— |
1280 |
— |
Баклан большой . . |
1630 |
— |
1250 |
1390 |
1320 |
— |
Нырок красноголовый |
1120 |
— |
1570 |
1330 |
500 |
— |
Кряква ...... |
630 |
— |
1250 |
214 |
395 |
320 |
Куры (белый леггорн) |
495 |
— |
1100 |
10 |
270 |
160 |
Действительно, у черноморского дельфина-белобочки определение концентрации мышечного гемоглобина в различных мышечных комплексах показывало, что минимальная концентрация мышечного гемоглобина имеется в сердечной мышце (1240 мг—%), вдвое больше в диафрагме (2800 мг—%), втрое больше в мышцах, приводящих в движение хвостовые лопасти (3600 мг—%). У таких водоплавающих птиц, как баклан, нырки, кряквы, концентрация мышечного гемоглобина в различных мышечных комплексах значительно ниже, чем у дельфинов, а у наземных птиц ниже, чем у водоплавающих.
Относительная разница между хорошими и плохими ныряльщиками среди птиц в концентрации мышечного гемоглобина в различных мышечных комплексах может быть продемонстрирована даже без непосредственного определения концентрации мышечного гемоглобина. {115} Если у подопытных птиц после перфузии (полного удаления крови из сосудистой системы) снять кожу, то окраска мышц тушки таких близких форм, как нырковые и кряковые утки, будет резко отлична. У кряковой утки мышцы светло-розового цвета, а у нырковых — темно-вишневого цвета. Если рядом с утками положить тушку курицы после перфузии, то грудные мышцы будут кремового цвета, выделяясь даже на фоне относительно бедных мышечным гемоглобином мышечных комплексов кряковой утки.
Однако знание уровня концентрации мышечного гемоглобина во всех мышечных комплексах изучаемого животного еще недостаточно для суждения о биологическом значении мышечного гемоглобина. Можно получить гораздо более полное представление, если определить вес этих мышечных комплексов, так как в этом случае можно будет определить общее количество мышечного гемоглобина в организме данного животного. Нами было произведено взвешивание всех мышечных комплексов дельфина-белобочки, наряду с определением концентрации мышечного гемоглобина.
Вес четырех мышечных тяжей, благодаря которым дельфин передвигается, составляет 56,5% веса всей мускулатуры животного (38,5% приходится на долю спинных тяжей и 18%—брюшных), или 15,2% веса тела. Масса этих четырех мышечных тяжей почти в три раза превышает вес всего скелета! Здесь так же, как и у наземных животных, для передвижения мобилизуются все ресурсы организма и в первую очередь более половины веса всех мышц. Общий вес всей мускулатуры дельфина составляет 30—35% веса тела. В общем, вес мускулатуры дельфина как вторичноводнсго животного меньше, чем вес мускулатуры наземных млекопитающих Выше говорилось, что у таких наземных млекопитающих, как северный олень или дикий баран архар, общий вес мускулатуры составляет около 50% веса тела, и только у домашних овец, подвижность которых сильно ограничена по сравнению с их дикими родичами, вес мускулатуры составляет 30—32% веса тела.
По-видимому, для плавания не последнюю роль играет подкожный жир, количество которого меняется у дельфина в зависимости от сезона. Жир вместе с кожей составляет свыше 40% веса тела. Если допустить, что {116} на долю кожи приходится 7—8% веса тела (у овец кожа с шерстью составляет 9—10% веса тела), то станет очевидным, что в основном энергия расходуется лишь на передвижение дельфина. При таком количестве жира его удельный вес будет близок удельному весу воды, го есть животное во взвешенном состоянии не будет затрачивать энергию на поддержание тяжести тела. Этим объясняется тот факт, что дельфин на передвижение расходует меньше энергии, чем его наземные собратья.
Как уже отмечалось выше, о затратах энергии косвенно можно судить по обеспеченности организма гемоглобином, который доставляет кислород работающим мышцам — основным потребителям кислорода. По нашим данным концентрация мышечного гемоглобина в мышце сердца дельфина примерно в три раз выше, чем у наземных животных, а в мышечных тяжах, приводящих организм дельфина в движение,— в 10 раз и более. Исходя из данных о концентрации мышечного гемоглобина в различных мышечных комплексах и весе этих комплексов, мы рассчитали, каким количеством мышечного гемоглобина располагает дельфин помимо гемоглобина крови.
Оказалось, что на 1 кг тела дельфина приходится около 11 г гемоглобина. А если суммировать вес гемоглобина в крови и мышцах, то на долю мышечного гемоглобина будет приходиться почти половина общего количества гемоглобина (табл. 5, 6), тогда как у наземных млекопитающих эта величина редко превышает 15% общего количества гемоглобинов. Все это означает, что мышцы дельфина являются одним из самых мощных резервов кислорода. Благодаря этому общая обеспеченность гемоглобином организма дельфина превышает 23 г на килограмм веса тела. Это большая величина, так как у большинства наземных млекопитающих обеспеченность гемоглобином значительно ниже: у домашней овцы не более 10 г гемоглобина на кг веса тела, у морской свинки менее 5 г и лишь у северного оленя эта величина поднимается до 20 г.
Если учесть, что у водных животных, обитающих в условиях, где действие сил гравитации резко ослаблено, имеются такие мощные очаги синтеза гемоглобина, то несомненно их можно рассматривать как одну из основных форм адаптации к «гипогравигационным» условиям {117} водной среды. Справедливость изложенных соображений в отношении форм адаптации дельфина можно проверить на другом представителе водных млекопитающих— тюлене, который хотя и не порвал полностью с сушей, но очень хорошо освоил и водную среду. Тюлени, как известно, более искусные ныряльщики, чем дельфины, ибо они могут быть под водой до 15 минут, т. е. значительно дольше, чем дельфины.
Исследования, проведенные на каспийском тюлене, полностью подтвердили эти предположения. Оказалось, что концентрация гемоглобина в крови тюленя также имеет рекордную величину — 22,6—23,0 г—%. Это значит, что кислородная емкость крови тюленя превышает 30 объемных % кислорода. Если учесть, что количество крови у тюленя составляет в среднем около 13% веса тела, то обеспеченность организма тюленя гемоглобином крови составит в среднем около 24 г на 1 кг веса тела. По-видимому, это рекордная величина обеспеченности организма гемоглобином крови, вдвое превышающая подобную величину у дельфина. Однако помимо гемоглобина крови тюлень обладает также огромным количеством мышечного гемоглобина. Даже после перфузии (а эта операция на тюлене проходит с большим успехом, чем у дельфина, так как у тюленя длинная шея и яремная вена, через которую осуществляется перфузия, легко доступна) мышцы тела имеют темно-вишневый цвет, что свидетельствует об очень высокой концентрации мышечного гемоглобина (таблица 5).
Так же, как и у дельфина, концентрация мышечного гемоглобина в различных мышечных комплексах тюленя
Таблица 5, Концентрация мышечного гемоглобина в различных мышцах каспийского тюленя (в мг—%)
Вес |
Пол |
Сердце |
Диаф- |
Задние |
Шея |
Передн. |
Туло- |
Мышечный |
18,5 |
♂ |
1790 |
4500 |
6375 |
— |
5925 |
9300 |
23,25 |
12,0 |
♀ |
1920 |
3180 |
— |
1890 |
— |
5100 |
15 62 |
16,9 |
♀ |
— |
2137 |
3075 |
1260 |
2588 |
3900 |
9,76 |
23,5 |
♀ |
1395 |
4425 |
5063 |
1560 |
4275 |
5500 |
14,68 |
25,0 |
♀ |
1328 |
2625 |
5025 |
1478 |
4650 |
6000 |
13,24 |
45,5 |
♀ |
1328 |
2700 |
4950 |
1470 |
3735 |
5400 |
14,37 |
| {118} |
неодинакова: наименьшая в сердечной мышце и мышцах шеи (1300—1500 мг—%), вдвое больше в диафрагме и в мышцах конечностей (4000—5000 мг—%), максимальная в мышцах туловища (до 6000 мг—%).
Однако эти данные о концентрации мышечного гемоглобина могут получить иное, более сильное «звучание», если определить вес всех перечисленных групп мышц. По данным, полученным в нашей лаборатории, общий вес поперечно-полосатой мускулатуры каспийского тюленя достигает 30% веса тела. В пересчете на килограмм веса тела количество мышечного гемоглобина тюленя составит в среднем около 15 г. А общее количество гемоглобина в организме тюленя в пересчете на единицу веса тела составит около 40 г на кг веса тела (таблица 6).
Таблица 6. Общее количество гемоглобина крови и мышц у некоторых млекопитающих
Гемоглобин, г/кг |
Общий |
Доля мы- |
||
крови |
мышц |
|||
Человек ..... |
12,8 |
0,57 |
13,4 |
3,5 |
Северный олень . . |
17,3 |
3,50 |
20,8 |
16,8 |
Морская свинка . . |
5,2 |
0,40 |
5,1 |
7,7 |
Дельфин-белобочка |
11,9 |
11,00 |
22,9 |
48,0 |
Тюлень каспийский |
24,2 |
15,00 |
39,4 |
38,6 |
В таблице даны рекордные цифры. Среди современных животных мы пока не знаем более высокого оснащения организма животного гемоглобином. Совершенно очевидно, что способность тюленя быть под водой около 15 минут определяется в первую очередь этим.
Обращает на себя внимание и еще одна особенность тюленя — огромное количество подкожного жира, достигающее у взрослых тюленей 50% веса тела. Помимо теплоизолирующей роли подкожный жир уменьшает удельный вес тела тюленя, вследствие чего тюлень находится в состоянии, близком к состоянию невесомости и практически тратит энергию лишь на передвижение в период пребывания в воде.
Экспериментальные данные о содержании гемоглобина в крови и мышцах, весе скелета и костного мозга {119} тюленя такого же порядка, что и у дельфина, хотя оснащенность гемоглобином крови и мышц у тюленя более высокая. Следовательно, данные, характеризующие степень приспособленности тюленя к водным условиям существования, подтверждают мысль о том, что относительный вес скелета позвоночных животных в основном определяется действием сил гравитации.
На суше, где действие сил гравитации проявляется в полной мере, у высших представителей наземных животных относительный вес скелета достигает 20% веса тела, тогда как у вторичноводных млекопитающих, обитающих в среде, где действие сил гравитации в сильной степени ослабляется, относительный вес скелета снижается до 5% веса тела. Трудно дать иное объяснение приведенным выше фактам из жизни вторичноводных животных.
Если предлагаемое объяснение фактов, касающихся особенностей структуры и функций этих структур у водных и наземных позвоночных животных, правильно, го интересно рассмотреть их и у других представителей млекопитающих и птиц, специализация которых к водному образу жизни не зашла так далеко, как у китообразных и ластоногих. Можно ожидать, что величины, характеризующие особенности крови, мышц, скелета, костного мозга, займут промежуточное положение между наземными и водными животными.
В этом плане из млекопитающих представляют интерес ондатра, нутрия, кутора, которые ведут полуводный образ жизни. Среди птиц в разной мере приспособлены к водному образу жизни утка и (речные и нырковые) баклан, чомга, пингвин. Из них, пожалуй, только пингвин наиболее приспособлен к водным условиям существования. Все остальные в меньшей степени адаптировались к жизни в воде. Однако ни одна из птиц не может конкурировать с китообразными и даже ластоногими, хотя среди них имеются прекрасные ныряльщики, не уступающие по длительности ныряния мелким китообразным (таблица 7).
Совершенно очевидно, что способность выключать внешнее дыхание на длительные сроки обусловлена соответствующими механизмами, с помощью которых создаются резервы кислорода, обусловливающие ту или иную длительность ныряния. Вместе с тем, адаптация {120} к водным условиям существования касается не только этих структур, но и структур, ответственных за передвижение в водных и наземных условиях. Ведь практически млекопитающим и птицам (за исключением китообразных) приходится адаптироваться и к наземным и к водным условиям. Естественно, происходит частичная специализация, которая позволяет вести такой земноводный образ жизни. Такая специализация касается особенностей кожных покровов, перепонок между пальцами конечностей, формы конечностей и др.
Что же касается сравнительных физиологических и биохимических исследований, то они, к сожалению, не получили еще должного размаха, и это сильно затрудняет решение эволюционных проблем. По-видимому, здесь играют роль две причины — трудность добывания нужного биологического материала и недостаточная осведомленность физиологов и биохимиков с зоологическими дисциплинами, такими, как зоогеография, экология и другие. Во всяком случае, пока имеется очень мало экспериментальных данных, характеризующих физиолого-биохимические аспекты адаптации этих двух групп животных к водным условиям существования. В таблице приводятся некоторые данные, характеризующие адаптационные механизмы водных птиц и млекопитающих.
У ондатры, нутрии и куторы скелет в два раза более тяжелый, чем у дельфина и тюленя. Оснащенность организма гемоглобином у ондатры ниже, однако достаточно высок уровень мышечного гемоглобина, который составляет почти 30% общего количества гемоглобина.
Данные, приведенные выше, характеризующие особенности крови, гемоглобина, очагов синтеза гемоглобина водных млекопитающих, можно рассматривать как убедительное доказательство справедливости выдвигаемого тезиса, согласно которому ослабление действия сил гравитации на организм позвоночных животных приводит к значительному изменению состояния очагов синтеза гемоглобина крови, о чем можно судить по изменению относительного веса скелета и его составной части костного мозга.
Приспособления, выявленные в организме водных млекопитающих, можно рассматривать как аргумент в обоснование тезиса, согласно которому длительное
| {121} |
Таблица 7. Количество крови, гемоглобина крови, костного мозга и мышечного гемоглобина у водных млекопитающих и птиц
Животное |
Вес, кг |
Ске- |
Костный мозг |
Кровь |
Гемоглобин, |
Мышечный |
||
тела |
скелета |
крови |
мышц |
|||||
Дельфин |
47,0 |
7,6 |
2,0 |
28,0 |
6,5 |
11,9 |
11,0 |
48,0% |
Тюлень |
49,0 |
5,0 |
2,0 |
27,0 |
13,0 |
24,2 |
15,2 |
38,6% |
Ондатра |
0,70 |
11,5 |
2,1 |
18,3 |
8,2 |
12,8 |
5,0 |
28,1% |
Кутора |
0,017 |
12,8 |
— |
— |
11,8 |
19,3 |
— |
19,6% |
Баклан-большой |
1,90 |
10,4 |
2,9 |
26,8 |
14,3 |
24,5 |
6,0 |
28,5% |
Чомга |
0,71 |
11,0 |
2,6 |
23,6 |
18,3 |
17,5 |
7,0 |
16,2% |
Утка кряква |
1,30 |
10,6 |
2,0 |
— |
9,9 |
8,3 |
1,6 |
|
Цесарка |
1,50 |
13,2 |
2,2 |
25,1 |
9,5 |
9,1 |
||
Куры (белый леггорн) |
1,64 |
11,5 |
2,0 |
22,0 |
6,5 |
4,6 |
0,9 |
16,3% |
пребывание в условиях невесомости должно привести к резкому нарушению деятельности очагов синтеза гемоглобина, в первую очередь скелета и костного мозга.
Таким образом, имеющиеся данные по особенностям биологии наземных позвоночных животных, с одной стороны, ,и водных млекопитающих, с другой, позволяют сделать вывод, что усиление биологического эффекта действия сил гравитации приводит к увеличению кроветворной функции скелета. Это свойственно позвоночным животным, обитающим на суше, о чем можно судить по увеличению относительного веса скелета и его органической составной части — костного мозга. Ослабление биологического эффекта действия сил гравитации в водных условиях приводит к ослаблению кроветворной функции скелета, наряду с усилением мощности очага синтеза гемоглобина в поперечно-полосатой мускулатуре тела водных млекопитающих. Именно данные об особенностях синтеза гемоглобина у наземных и водных млекопитающих дают основание для прогноза реакций организма в условиях длительной невесомости, в первую очередь в деятельности такой жизненно важной
| {122} |
Среда |
Суша |
Вода |
Космос |
Характер дей- |
Максимальное |
Резко ослабленное |
Практически полное от- |
Возможность |
Поведение организма на суше и в во- |
Современная техника пока |
|
Результаты |
Резкое усиление |
Ослабление гемопоэ- |
? |
структуры, как скелет, биологическое значение которого до сих пор еще недооценивается многими биологами.
Если суммировать имеющиеся данные по действию сил гравитации на организм, в первую очередь на организм позвоночных животных, главным образом млекопитающих, водных и наземных, то все они могут быть представлены в виде схемы.
| {123} |
Глава V
Современная техника пока не дает возможности создать на Земле условия, лишенные действия сил гравитации. Следовательно, пока нельзя поставить опыты, которые дали бы прямой ответ на вопрос о том, как влияет на организм состояние длительной невесомости. Вместе с тем этот вопрос далеко не праздный, ибо за освоением нашего ближайшего спутника — Луны встанет вопрос о полетах на ближайшие планеты — Марс и Венеру. Нет никаких оснований думать, что человек ограничит себя только этими целями.
Но независимо от темпов освоения ближайшего межпланетного пространства, современная биология и медицина должны уже сейчас дать ответ на вопрос: что представляет собой жизнь без гравитации, или состояние длительной невесомости? Что это за фактор? Полезен он для организма, безразличен или чреват вредными последствиями?
В зависимости от ответа на этот вопрос может возникнуть проблема профилактики, решить которую можно будет только на основании изучения природы длительной невесомости, жизни без гравитации.
При рассмотрении любых аспектов проблемы невесомости следует исходить из того бесспорного положения, что жизнь на нашей планете возникла и развивалась в условиях гравитационного поля. Значит, все структуры организма адаптированы к этим условиям и коррелятивно связаны при функционировании с другими структурными образованиями организма. Если исходить из признания принципа единства организма и среды, то силы гравитации нельзя игнорировать, ибо организмы, возникшие и развивающиеся в гравитационном {124} поле, имеют соответствующий настрой функционирования в этом поле.
Исходя из этих положений следует считать, что пребывание в состоянии невесомости, например при полете на Марс, который будет длиться около 7 месяцев только в один конец, означает переход организма в другую среду обитания. Многие из компонентов прежней среды обитания будут сохранены в том замкнутом экологическом комплексе, который будет создан в космическом корабле. Но тем не менее, известно, что даже частичное изменение парциального давления кислорода вызывает сдвиг в ходе жизненных процессов, в первую очередь в органах аэрации и гемодинамики. Если же организм переходит в другое гравитационное поле, то он должен полностью адаптироваться к нему. Так было при освоении суши и при вторичном освоении воды, так должно быть при переходе в состояние длительной невесомости. Разница лишь в том, что переход в состояние невесомости является чистым опытом, в котором будет четко выявлена роль невесомости. Полагать, что устранение такого мощного фактора, как силы гравитации, останется не замеченным организмом, значит допустить ошибку, возможно чреватую многими весьма нежелательным последствиями.
Если, однако, переходить от общих положений о вероятности воздействия на организм длительной невесомости, к конкретным проявлениям этого воздействия, то необходимо искать аналогии в тех формах адаптации организма, которые имели место при переходе от наземного к водному образу жизни у различных представителей позвоночных животных, а также при появлении млекопитающих. Детальнейший их анализ должен дать много для решения проблемы невесомости.
При переходе в состояние невесомости наиболее резким изменениям должен подвергнуться общий уровень энергетики организма. В этом плане анализ особенностей энергетики у вторичноводных животных и у плодов млекопитающих при внутриутробном развитии даст весьма ценные материалы.
Ранее отмечалось, что проблема невесомости — не только проблема чисто небесная, связанная с освоением космических просторов, она в то же самое время проблема чисто земная, решение которой важно для {125} фундаментальных земных проблем. Справедливость этого тезиса подтверждает анализ фактов из жизни некоторых представителей наземных животных, в частности птиц и млекопитающих.
Известно, что все предки сельскохозяйственных животных обладали более совершенными механизмами преодоления сил гравитации, так как были прекрасными скороходами (дикие лошади, бараны и другие). Их потомки — наши домашние животные — уже в большой степени утратили эти механизмы. Дикий баран архар развивает скорость свыше 60 км в час, а домашние овцы способны лишь к весьма ограниченным передвижениям.
По существу домашние условия — новая среда обитания, резко отличная от естественной среды обитания предков наших сельскохозяйственных животных или их диких сородичей. Адаптация к этой новой среде вызвала глубокие изменения как в поведении, так и в строении организма, в первую очередь в органах передвижения как основных структур преодоления сил гравитации. Дикое животное все время находится в состоянии напряжения, готовности к прыжку, к бегу. Одомашненные животные в значительной мере потеряли эту «бдительность», что, несомненно, отражается на всей энергетике организма. Усилия, которые приходится делать диким животным в случае опасности или других неблагоприятных условий, у домашних животных значительно ослаблены.
На первый взгляд изменение природы организма дикого животного кажется положительным. Однако в результате сильно уменьшается вес мускулатуры и скелета, т. е. уменьшается вес структур, определяющих не только благополучие организма, но и имеющих высокую продуктивную ценность. В частности, у лося, северного оленя и архара вес мускулатуры по отношению к весу тела составляет около 50% веса тела, а у домашних овец, например у породы «Советский меринос», вес мускулатуры составляет лишь немногим более 30% веса тела. Правда, у домашних животных создаются такие резервы жира, каких у их диких родичей обычно нет.
| {126} |
При сравнении сельскохозяйственных животных и их диких сородичей представляет интерес поведение самок в период беременности, особенно в случае многоплодных животных. В это время на организм матери резко возрастает дополнительная нагрузка, вследствие чего возрастают энергетические затраты на питание плода и на преодоление сил гравитации. У многоплодных романовских овец вес тела к концу беременности возрастает на 22,5% по сравнению с исходным весом, а в абсолютных цифрах до 17 кг (в случае четверни); в случае одинцов — только на 10% веса, или 6,5 кг.
Мы не имеем данных о весе самок диких овец в период беременности. Поэтому представляют интерес живородящие водные животные: китообразные из млекопитающих, и катраны, и скаты из хрящевых рыб. В силу специфических особенностей водного образа жизни у китообразных рождается главным образом по одному детенышу, но к моменту рождения они достигают очень крупных размеров: половины длины тела матери и до 10% веса ее тела. Однако в водной среде действие сил гравитации резко ослаблено, и самке носить плод легко по сравнению с наземными условиями.
Иное дело наземные условия, где действие сил гравитации проявляется в полной мере. В этих условиях о размерах энергетических затрат можно судить по интенсивности потребления кислорода в период беременности и в период яловости. Имеется очень мало данных, характеризующих этот ответственный период с физиологической стороны. Они свидетельствуют о том, что у свиней в первой половине беременности повышение энергетических затрат достигает 30% по сравнению с исходным состоянием, а во второй половине — 50%.
Однако у овец в первой половине беременности уровень энергетических затрат мало отличается от исходного, а во второй половине повышается до 17%. У кроликов в первой половине беременности энергетические затраты превышают исходный уровень в среднем на 10%, а во второй половине — на 13%, в конце ее уровень обмена веществ значительно повышается. В опыте, проведенном в Узбекистане, каракульские {127} одноплодные матки потребляли кислорода за 5—10 дней до скота на 30% больше по сравнению с яловыми, а матки с двумя плодами — на 42%. Аналогичные результаты были получены у тонкорунных овец. Романовские овцы с одним и двумя плодами на втором-третьем месяце беременности потребляли кислорода больше, чем яловые, на 6—8%, матки с тремя — 18%, а в конце ее соответственно на 24, 51 и 48% больше.
Следует отметить также, что энергетические затраты у разных видев сельскохозяйственных животных в период беременности неодинаковы. По-видимому, эта разница определяется не только различиями в образе жизни, но и плодовитостью (многоплодное или одно-плодное животное).
Обращает на себя внимание и то обстоятельство, что уровень энергетических затрат в период беременности меньше, чем в период лактации. Казалось бы, что период беременности настолько ответственное время в жизни вида, сопряженное со столь глубокой перестройкой организма матери, обеспечивающей исключительно высокие темпы развития плода, что он потребует во много раз больше энергии, чем в период лактации. Однако факты показывают, что это не так. Понять эти особенности можно будет лишь в случае дифференцированного учета энергетических затрат не только организма матери, но и организма плода.
В целом же в период беременности, особенно в его последний этап, вес тела матери значительно увеличивается по сравнению с исходным весом, в связи с чем размеры энергетических затрат весьма значительны.
О масштабах увеличения веса тела матери в период беременности у различных представителей млекопитающих можно судить по данным, представленным в таблице 8 (данные Лейч, Хиттен, Билевич, 1959). Они свидетельствуют о значительных вариациях веса тела матери в период беременности у разных представителей млекопитающих. Рекордсменами являются некоторые представители грызунов, например морская свинка, общий вес новорожденных которой достигает 63%, т. е. больше половины веса тела матери. На втором месте обыкновенная {128} полевка — общий вес новорожденных составляет 50% веса тела матери. У сельскохозяйственных животных— свиньи, коровы, лошади — вес новорожденного в среднем равен 7,5% веса тела матери, у хищников — 12%. Все эти данные хорошо объясняются образом жизни животных, их отношением к гравитационному полю. У морской свинки, ведущей почти неподвижный образ жизни, увеличивается вес тела на 60%, тогда как у копытных и хищников только на 7—15%.
Несколько неожиданны данные то весу тела новорожденных у рукокрылых, вес которых колеблется от 20 до 35% веса тела матери. Если учесть, что это летающие млекопитающие, то можно только поражаться превосходным летным качествам этих животных, которые в период беременности в состоянии выдерживать такую нагрузку. Размеры этой нагрузки, по-видимому, еще более значительны, если учесть, что в период беременности помимо плода имеются еще околоплодные жидкости.
Это обстоятельство обычно не учитывалось, ибо считалось, что дополнительная нагрузка определяется лишь самим плодом, его потребностями. Изучение динамики изменения веса эмбрионов мериносовых овец свидетельствует о весьма бурных темпах роста эмбриона на первых этапах развития, затухающих к концу внутриутробного развития: в течение первых 30 дней вес тела эмбриона увеличивается в 2500 раз (если оплодотворенное яйцо весит 1 мг), во второй месяц в 23,2 раза, в два последних месяца — лишь в 2,75 раза.
Имеющиеся данные по динамике и изменению веса эмбрионов овец показывают, что в последние два месяца беременности плод накапливает основную массу своего тела, равную 85% веса при рождении. Это обстоятельство дало повод некоторым зоотехникам утверждать, что решающим условием получения здорового приплода и сохранения нормального состояния материнского организма является усиленнее кормление матери лишь в последней стадии беременности. Считалось, что усиленное и полноценное кормление овец во второй половине беременности обеспечивает также накопление значительных резервов в организме, крайне необходимых при последующей лактации.
Вывод этот, особенно в первой части, безусловно спорный, так как не учитываются начальные стадии
| {129} |
Таблица 8. Вес организма матери и новорожденного у млекопитающих
Вид |
Число |
Вес |
Вес ново- |
||
матери |
новорож- |
всех |
|||
в г |
|||||
Рукокрылые |
|||||
Летучая мышь |
1 |
19,7 |
6,8 |
6,8 |
34,5 |
Летучая мышь |
1 |
7,2 |
2,08 |
2,98 |
28,9 |
Насекомоядные |
|||||
Еж |
7 |
725 |
13,9 |
98 |
13,5 |
Землеройка |
5 |
16,6 |
0,8 |
4,0 |
24,1 |
Грызуны |
|||||
Домашний кролик |
9 |
3200 |
81 |
729 |
22,8 |
Заяц-русак |
3 |
3000 |
132 |
370 |
12,6 |
Белка |
3,2 |
600 |
15 |
48,5 |
8,1 |
Обыкновенная полевка |
8,5 |
31 |
1,85 |
15,6 |
50,3 |
Морская свинка |
3,7 |
560 |
95 |
352 |
63,0 |
в кг |
|||||
Приматы |
|||||
Человек |
1 |
56 |
3,2 |
3,2 |
5,7 |
Горилла |
1 |
60 |
1,75 |
1,75 |
3,0 |
Ластоногие |
|||||
Гренландский тюлень |
1 |
120 |
11,8 |
11,8 |
9,8 |
Хищники |
|||||
Полярный медведь |
1,5 |
280 |
0,59 |
0,89 |
0,3 |
Лисица |
6,8 |
4,200 |
0,1 |
0,680 |
16,2 |
Копытные |
|||||
Лошадь |
1 |
800 |
71 |
71 |
8,9 |
Свинья |
8,3 |
163 |
1,14 |
9,5 |
5,8 |
Северный олень |
1 |
105 |
5,6 |
5,6 |
5,3 |
Овца |
2 |
52,3 |
3,65 |
7,3 |
14,0 |
Неполнозубые |
|||||
Муравьед |
1 |
28 |
1,48 |
1,48 |
5,4 |
в т |
|||||
Китообразные |
|||||
Фиивал |
1 |
50 |
1,5 |
1,5 |
3,0 |
| {130} |
исключительно бурного роста эмбриона, которые обусловливаются соответствующей подготовкой организма матери. Как справедливо заметил английский физиолог Баркрофт, «сцена готовится прежде, чем начнется спектакль». Это значит, что уровень и качество питания в начальной стадии беременности — не менее решающее условие, ибо здесь закладываются основы будущего благополучия организма плода, а не только в последние два месяца, когда большие объемы пищи едва ли могут быть эффективно использованы.
Таким образом, у многоплодных животных в период беременности на организм матери ложится большая дополнительная нагрузка, достигающая пятой части веса тела. Естественно, что возрастают и энергетические затраты, связанные не только с питанием плода, но и с необходимостью преодоления сил гравитации.
У птиц с их внеутробным типом развития такой большой перестройки организма матери, которая характерна для млекопитающих не происходит. Правда, организм матери несет основную нагрузку в период яйцекладки, снабжая каждое яйцо «сухим пайком» на весь период развития вплоть до выклева. И во многих случаях нагрузка достигает весьма внушительных размеров. Однако в период яйцекладки организм матери птиц не знает увеличения воздействия сил гравитации на организм. Это — удел млекопитающих. На стр. 132 приводятся некоторые данные по весу отдельных яиц и яйцекладок некоторых видов птиц.
Как видно из приводимых цифр, у разных видов птиц нагрузка на организм матери различна и колеблется от 5% веса тела (кондор) до 160% (славка). У бескилевых птиц эти величины составляют 20—36% веса тела, у гуся 60%, У кур около 55%, но в некоторых случаях до 450% веса тела (200 яиц в год).
Совершенно очевидно, что, если развитие эмбриона протекает вне организма матери, проблема возрастания усилий по преодолению сил гравитации отсутствует, хотя материнский организм птиц во многих случах несет
| {131} |
Вес |
Вес |
Вес яй- |
Вес яиц |
|
Бескилевые |
||||
Страус Найду.......... |
20,0 |
575 |
11 |
36 |
Киви.............. |
2,5 |
455 |
1 |
20 |
Килевые |
||||
Цапля............. |
2,6 |
95 |
4 |
15 |
Кондор............. |
11,0 |
275 |
2 |
5 |
Курица домашняя........ |
2,5 |
55 |
25 |
55 |
Глухарь ............ |
2,5 |
50 |
8 |
16 |
Колибри ............ |
3,2 |
0,45 |
2 |
25 |
Крапивник ........... |
9,5 |
1,3 |
7 |
95 |
Синица............. |
11,0 |
1,3 |
10 |
118 |
Славка............. |
5,0 |
0,72 |
11 |
158 |
весьма большую нагрузку, синтезируя огромное количество резервных веществ, необходимых для развития эмбриона, которые в ряде случаев достигают веса тела матери, а у домашних кур превышает вес тела матери в три-четыре раза. Такая задача, видимо, непосильна для организма млекопитающих, поэтому у них все сильно растянуто во времени. Если у птиц самый большой срок инкубации около двух месяцев (страусы), то у млекопитающих сроки внутриутробного развития иногда растягиваются до года и более (слоны, жирафы).
По-видимому, организм птиц обладает большими возможностями биосинтеза пластических веществ, чем организм млекопитающих, и не потому, что клетки тела птиц обладают какими-то особыми свойствами, а вследствие разных типов развития. Внутриутробный тип развития— более совершенная форма, поскольку его ранние стадии протекают в менее уязвимых условиях, тогда как внеутробный тип развития сопряжен с большим риском.
Млекопитающие не могут соревноваться с птицами по количеству приплода. Даже более ограниченные возможности млекопитающих в численности приплода создают дополнительные трудности в преодолении сил гравитации, которые не испытывают птицы. По этой причине живой вес млекопитающих в значительной степени меняется, вес же птиц не подвержен изменениям. {132}
Проблема внутриутробного развития имеет и другой, не менее важный аспект — энергетический уровень материнского организма и плода в период беременности. Но даже данные по суммарной энергетике материнского организма в период беременности (если мы будем ими располагать) не дадут представления о распределении энергии между массой материнского организма и плода. Можно пока предполагать, что эмбриональные ткани плода будут расходовать относительно больше энергии, чем ткани материнского организма. Известно, что в онтогенезе животного ранние стадии развития характеризуются более интенсивным метаболизмом, чем поздние стадии.
Справедливость этого предположения как будто бы подтверждается данными по обеспеченности организма плода гемоглобином. Организм плода, особенно к моменту рождения, обладает большим количеством гемоглобина, превышающим в два-три раза количество гемоглобина в организме матери в пересчете на единицу веса тела. Пропорционально количеству гемоглобина плод обладает также и более высоким содержанием костного мозга (очага синтеза гемоглобина) и относительно более тяжелым весом скелета, чем организм матери. Это явление (гетерохрония) в развитии скелета плода млекопитающих, впервые открытое В. Я. Броваром, получило свое объяснение лишь в связи с количественной характеристикой крови и гемоглобина в организме плода. Данные по обеспеченности организма плода гемоглобином косвенным образом подтверждают справедливость предположения о более высоком уровне метаболизма в организме плода по сравнению с материнским организмом и свидетельствуют о необоснованности представления об угрозе кислородного голодания в период внутриутробного развития. Однако прямые определения интенсивности потребления кислорода организмом матери и плода в период беременности показали, что организм плода потребляет кислорода примерно в три-четыре раза меньше, чем новорожденное животное. Это было совершенно непонятно. {133}
Баркрофт был первым исследователем, который установил, что плод козы потребляет неожиданно мало кислорода, примерно в три-четыре раза меньше, чем новорожденные козлята. Эти данные были настолько неожиданно малыми, что сам он позднее относился к ним критически. Другие исследователи, изучавшие этот вопрос, подтвердили, что интенсивность потребления кислорода организмом плода ниже, чем у новорожденных животных (табл. 9).
Таблица 9. Потребление кислорода плодом и новорожденным
Возраст плода, дни |
Вес плода, кг |
Потребление |
79-87; 85 |
0,32—0,52 |
4,0 |
117—138; 125 |
2,13—5,23 |
3,7 |
136—144; 140 |
3,48—6,03 |
4,5 |
Новорожденный козленок |
3,5 |
16,0 |
Такие же величины потребления кислорода были получены на новорожденных телятах, жеребятах, поросятах.
Обсуждая полученные результаты, Ачесон и другие ученые отмечали, что у ягнят в возрасте более 12 часов (после рождения) «имеет место резкое повышение потребления кислорода, которое требует дальнейших исследований». Баркрофт, впервые открывший этот феномен, писал, что важнейшей причиной резкого усиления потребления кислорода новорожденного является мышечный тонус, исчезающий при помещении новорожденного в теплый солевой раствор, и снова появляющийся, когда животное просто находится в «покое».
О более низком уровне окислительных процессов в клетках тела свидетельствовали данные по определению активности окислительных ферментов у лошади. Согласно им, активность цитохромоксидазы у плода за 6 недель до рождения составляла лишь 56% активности взрослой лошади, сукциноксидаза и сукциндегидрогеназа соответственно — 31 и 29%. Эта более низкая активность ферментов рассматривалась также как адаптация к условиям ограниченного поступления кислорода. Об этом же свидетельствуют данные И. А. Аршавского: у только что родившегося человека в крови содержится 4—5 млн. {134} эритроцитов и 70—80% гемоглобина (определяемого методом Сали), а через 1-2 дня после рождения уровень эритроцитов повышается до 6—7 млн. и гемоглобина до 110-145%.
Основные выводы, которые делались на основании полученных данных, сводились к тому, что энергетика тканей организма плода имеет более низкий уровень вследствие недостаточного поступления кислорода. В момент рождения энергетика организма переключается на более эффективный путь аэробного дыхания, а тканевые катализаторы увеличивают свою активность под влиянием резко возросшего напряжения кислорода в среде и охлаждения тела. На этом основании говорят о кислородном и температурном ударе, который получает плод в момент рождения. Это те стимулы, которые вызывают необходимые сдвиги в переходе на аэробный тип дыхания (имеется в виду, что в период внутриутробного развития преобладает анаэробный тип дыхания).
Эти объяснения носят, несомненно, надуманный характер и не соответствуют действительности. Известно, что гемоглобин плода имеет более высокое сродство к кислороду, чем гемоглобин материнского организма. Совершенно очевидно, что у птиц проблема обеспечения организма эмбриона кислородом решена весьма эффективно, ибо у них кислород поступает через поры скорлупы яйца и, следовательно, в этом случае нет оснований говорить об анаэробном дыхании. Нет оснований говорить и об анаэробном типе дыхания в период внутриутробного развития млекопитающих, так как они обладают во много раз более мощными механизмами обеспечения организма плода кислородом, чем это свойственно птицам. Об этом говорят представленные ниже данные о количестве гемоглобина в организме птиц и млекопитающих (в г на 1 кг веса тела):
Гуси | Утки | Цесарки | Овцы | Кролики | |
Новорожденные...... | 4,4 | 2,0 | 2,8 | 22,0 | 15,0 |
Взрослые......... | 8,5 | 6,4 | 7,2 | 8,0 | 8,0 |
Если у только что выклюнувшихся птенцов содержится в два раза меньше гемоглобина, чем у взрослых, то у млекопитающих организм новорожденного содержит {135} почти в два-три раза больше гемоглобина, чем материнский организм. Если у плода млекопитающих имеет место анаэробный тип дыхания, то чем объяснить такое огромное количество гемоглобина в пересчете на единицу веса тела? Именно поэтому рассуждения о кислородном ударе в момент рождения не имеют никаких оснований. Более того, говоря об анаэробном типе дыхания плода млекопитающих в период внутриутробного развития, допускают как бесспорное положение адекватность условий внутриутробных и условий после рождения (за исключением будто бы кислорода и температуры). Суть же вопроса состоит в том, что внутриутробные условия развития не адекватны наземным условиям существования и вовсе не потому, что новорожденного ожидает кислородный и температурный удар, а потому, что в период внутриутробного развития плод находится в водной среде во взвешенном состоянии, близком к состоянию невесомости, т. е. в среде, где действие сил гравитации резко ослаблено. Новорожденное млекопитающее попадает в условия, где действие сил гравитации проявляется в полной мере. Вот это-то последнее обстоятельство и является, с нашей точки зрения, той основной причиной, которая обусловливает резкое повышение интенсивности потребления кислорода в три-четыре раза, как это было установлено Баркрофтом и сотрудниками в 40-х годах и позднее другими исследователями.
Мы располагаем сейчас данными, подтверждающими справедливость этого положения. Водные животные, в частности рыбы, обладают менее интенсивным энергетическим обменом по сравнению с наземными позвоночными животными. Это происходит не потому, что они являются представителями низших позвоночных, а потому, что они обитают в «гипогравитационных» условиях, требующих меньших затрат энергии. Наземные животные тратят больше энергии именно на преодоление сил гравитации. В качестве важнейшего средства преодоления сил гравитации возник новый очаг синтеза гемоглобина в виде костного мозга, вследствие чего костный мозг можно {136} рассматривать как достояние наземных позвоночных животных.
Костный мозг достиг мощного развития у высших представителей позвоночных животных — у птиц и особенно у млекопитающих, составляя иногда свыше 10% веса тела животного. С этой точки зрения переход в «гипогравитационные» условия должен привести к обратному результату — уменьшению относительного веса скелета, а соответственно и костного мозга. Именно такого рода данные были получены нами при изучении очагов синтеза гемоглобина у водных млекопитающих— дельфинов и тюленей. Вес скелета у дельфинов и тюленей составляет в среднем 5—6% веса тела, т. е. почти в два раза меньше, чем у наземных млекопитающих (овцы, коровы, лошади). Скелет как основная система органов преодоления сил гравитации в новых условиях не получал должной нагрузки и стал редуцироваться вместе с костным мозгом, ибо потребность в энергии резко снизилась.
Если проводить параллели, то и плод в период внутриутробного развития, находясь в водной среде, должен иметь значительно менее интенсивный метаболизм, чем в наземных условиях. Соответствующие эксперименты Баркрофта и других ученых полностью это подтверждают. Именно момент перехода от внутриутробного к внеутробному развитию сопровождается повышением интенсивности метаболизма в три-четыре раза с 4—5 до 15—20 см3/кг/минуту.
Это интерпретация фактов, однако, содержит в себе одно противоречие. Ведь у водных млекопитающих ослабление интенсивности метаболизма в водных условиях сопровождалось снижением относительного веса скелета и костного мозга, тогда как. у плодов млекопитающих к концу внутриутробного развития и у новорожденных животных относительный вес скелета и костного мозга достигает рекордно больших величин— 20—22% веса тела для скелета и 13% для костного мозга, т. е. в два-три раза больше, чем в материнском организме.
В действительности здесь нет никакого противоречия, так как скелет в период внутриутробного развития не выполняет своей основной опорной функции, характерной для наземных условий, поскольку тело плода взвешено {137} в околоплодных жидкостях и, следовательно, находится в состоянии, близком к невесомости. В данной ситуации скелет уподобляется органам кроветворения первичноводных — селезенке, почкам, печени — и как полостной орган, не несущий опорной функции, достигает громадных размеров, превышая относительный вес скелета материнского организма в два-три раза.
Последнее обстоятельство очень важно, ибо проблема бесперебойного снабжения кислородом в период внутриутробного развития имеет исключительное значение. Основной путь поступления кислорода в организм плода — это контакт капилляров стенки матки с капиллярами плаценты. Естественно, что организм плода должен иметь определенные резервы кислорода и, следовательно, значительные количества гемоглобина, который синтезируется костным скелетом.
В этом случае особенности скелета определяются не силами гравитации, а теми потребностями в кислороде, которые на более ранних этапах развития удовлетворяются за счет гемопоэтической функции других структур организма плода (стенкой желточного мешка, селезенкой, почками или печенью). Таким образом, помимо общетеоретических обобщений о роли сил гравитации в эволюции наземных животных, которые можно сделать на основании анализа экспериментальных данных, имеется несколько обобщений, представляющих интерес в зоотехническом плане. Одомашнение, связанное с резким изменением поведения животного, его активности, ведет к ослаблению механизмов преодоления сил гравитации, в первую очередь костно-мышечных систем, т. е. к редукции ценнейшего компонента в продукции животноводства, взамен которого получает мощное развитие жировые депо.
Птицы, как группа позвоночных с внеутробным типом развития, обладают большими возможностями по сравнению с млекопитающими для продуцирования пластических веществ, используемых в период яйцекладки, часто превышающих вес тела материнского организма. Благодаря этому силы гравитации у них не являются фактором, лимитирующим возможности организма матери, как это имеет место у млекопитающих.
Вскрытие физиолого-биохимических особенностей внутриутробного развития плода млекопитающих, {138} связанных с низким уровнем энергетики вследствие пребывания в «гипогравитационных» условиях (по сравнению с наземными условиями существования), может быть использовано при разработке рационов животных в период беременности. В частности, мнение, согласно которому усиленное кормление необходимо лишь во второй половине беременности, не может рассматриваться как убедительное, ибо оно в такой же, если не в большей мере, необходимо в первой половине беременности, когда создаются условия для нормального развития плода, определяющего благополучие будущего животного, а тем самым его племенные и продуктивные возможности.
Рассмотренные выше примеры, касающиеся поведения организма в водных условиях (в случае вторично-водных млекопитающих, в период внутриутробного развития), свидетельствуют о резком изменении уровня обмена веществ по сравнению с наземными условиями существования. Как уже неоднократно говорилось, основная причина, обусловливающая эти особенности обмена веществ,— высокая (плотность воды, «снижающая» действие сил гравитации, которое в полной мере проявляется у наземных животных. Практически водные организмы взвешены в воде, и траты энергии, если животное в покое, минимальны. Как у рыб, так и у водных млекопитающих имеются механизмы, с помощью которых они могут менять свой удельный вес. Даже такие гиганты морей, как крупные китообразные, могут менять свой удельный вес, который у них почти равен удельному весу воды. Чтобы погрузиться в воду, водные млекопитающие ныряют на полувыдохе, так как на полном вдохе им трудно нырять (удельный вес в этом случае равен удельному весу воды или меньше).
Водная среда породила настоящих гигантов — синего кита, достигающего 150 т. Однако стоит такому гиганту, легко передвигающемуся в водной стихии на больших скоростях, попасть в наземные условия, как все кончается весьма трагически. В то же время среди наземных животных к категории гигантов могут быть отнесены лишь африканские слоны (крупные самцы, вес которых достигает 4,5 т) и бегемоты (вес которых колеблется от 2,5 до 4 т). Однако бегемоты ведут полуводный образ жизни, т. е. в этом случае вода, благодаря своей высокой плотности, облегчает вес тела животного. {139}
Представляют интерес расчеты американских палеонтологов в весе тела гигантских рептилий мезозоя. На основании определения объема реконструированных моделей динозавров был рассчитан вес, который оказался равным для тиранозавра 7 т, диплодока — 10 т, бронтозавра — 30 т и брахиозавра — более 70 т. Совершенно очевидно, что эти гиганты вели земноводный образ жизни, так как передвигаться по суше, где действие сил гравитации проявляется в полной мере, они не могли.
Впрочем предположение об образе жизни гигантских рептилий мезозоя верно будет лишь при одном условии, что гравитационное поле нашей планеты не изменялось на протяжении всей истории существования Земли. Недавно было высказано мнение, согласно которому гравитационное поле Земли не постоянно. Мысль, что гравитационное поле Земли менялось на протяжении геологического времени, была высказана в 1962 г. Б. В. Нейманом1 и в 1969 г. Л. С. Смирновым и Ю. Н. Любиной2.
Если это мнение подтвердится, то будет решен вопрос о причинах массовой гибели гигантских рептилий мезозоя. Резкое увеличение гравитационного поля Земли могло бы стать причиной их гибели. Но это пока лишь гипотеза, которую надо обосновать.
Все эти данные, которые характеризуют особенности организмов, обитающих на суше и в водных условиях, позволяют высказать прогноз об особенностях организма в условиях отсутствия гравитационного поля или в условиях длительной невесомости. (Физики утверждают, что гравитационное поле полностью не исчезает, но сила его действия может быть сведена к минимуму.)
Основные данные, на основании которых строится этот прогноз,— данные об особенностях рыб как первичноводных животных, наземных позвоночных животных и вторичноводных млекопитающих. Суть их сводится к тому, что органы движения, на которые падает вся {140} тяжесть в связи с необходимостью преодоления сил гравитации, претерпевают сильные изменения в процессе адаптации к пребыванию на суше и в воде. Сравнительный анализ весовых особенностей различных структурных образований организма некоторых представителей позвоночных животных при выяснении причин, обусловивших увеличение количества крови и гемоглобина у первых выходцев на сушу — амфибий, показал, что максимальные изменения претерпели мышечная система и скелет.
Накопленный с тех пор материал подтвердил справедливость положения, согласно которому в филогенезе позвоночных животных имеют место увеличение относительного веса скелета и продуктивных возможностей скелета, что находит свое выражение в массе крови и гемоглобина, содержащихся в сосудистой системе животных. В связи с тем, что размеры позвоночных животных несоизмеримы, предложен способ унификации в виде пересчета массы крови и гемоглобина на единицу веса тела. Это необходимо хотя бы потому, что даже среди одной группы млекопитающих имеются представители, вес которых достигает 150 т (синий кит) и 2—3 г (землеройка).
Ниже представлены средние данные веса скелета, количества крови и гемоглобина в организме для основных групп позвоночных животных. Они могут служить убедительным заключением всему сказанному выше об адаптационных изменениях организма животных в процессе эволюции позвоночных животных:
Эти данные показывают, что у наземных позвоночных в результате адаптационных изменений более тяжелый относительный вес скелета, примерно вдвое большая масса крови и почти десятикратное увеличение количества гемоглобина в пересчете на единицу веса тела.
Число видов |
Скелет, % |
Кровь, % |
Гемоглобин, г/кг |
|
Хрящевые.......... |
3 |
7,0 |
2,0 |
1,1 |
Костистые.......... |
8 |
8,5 |
2,9 |
1,8 |
Амфибии........... |
4 |
11,4 |
5,6 |
3,6 |
Рептилии .......... |
5 |
14,0 |
5,8 |
3,8 |
Птицы............ |
6 |
14,3 |
9,7 |
10,2 |
Млекопитающие....... |
18 |
14,2 |
7,3 |
12,1 |
| {141} |
Мы рассматриваем это увеличение веса скелета и массы крови как следствие адаптации к новым условиям, где действие сил гравитации проявляется в полной мере, или как прямое влияние сил гравитации на организм позвоночных животных.
Если верны эти рассуждения, то можно ожидать в случае перехода позвоночных животных в среду с ослабленным действием сил гравитации уменьшения относительного веса скелета. Изучение особенностей некоторых представителей водных млекопитающих (дельфин, тюлень) полностью подтвердило наш прогноз. Эти два обстоятельства (появление наземных позвоночных животных и вторичноводных млекопитающих) и дали нам возможность высказать мысль, что переход животного в состояние длительной невесомости должен привести к нарушению нормальной деятельности скелета, как гемо-поэтической системы.
Жизнь на нашей планете развивалась в условиях гравитационного поля определенной интенсивности. Устранение гравитационного поля в условиях длительных космических полетов не может остаться «не замеченным» организмом космонавтов. Причем исчезновение гравитационного поля никак нельзя рассматривать как фактор полезный или безразличный для организма. Это такой же грозный фактор, как и космическая радиация.
В литературе иногда встречаются утверждения, что длительная невесомость достаточно безобидный фактор, к которому в конце концов можно привыкнуть. Об этом пишет в частности английский физик и писатель Артур Кларк в своей книге «Черты будущего». Он говорит о том, что из всех сил природы — сила тяготения наиболее таинственная и наиболее беспощадная, что она властвует над нами от рождения до самой смерти, убивая или увеча за каждый промах1. Далее Кларк пишет, что в будущем человек освободится от сил тяжести, состояние невесомости будет естественным явлением. Б будущем невесомость, или ослабленная сила тяжести, станет привычным и, может быть, даже нормальным {142} состоянием. «Вероятно, придет и такое время,— пишет он далее,— когда на космических станциях и планетах с малой силой тяжести будет жить больше людей, чем на Земле, а когда будет написана история человечества, то примерно сто миллиардов людей, которые уже прожили свои полные трудностей жизни, борясь против гравитации, окажутся лишь ничтожным меньшинством... Очень может быть также, что наши потомки, освоившие Космос, будут столь безразличны к силе тяжести, как и наши отдаленные предки, которые без особых усилий плавали в море.
Ведь даже теперь большинство живых существ на нашей планете не ощущает силы тяготения. Правда, оно господствует лад крупными наземными животными — слонами, собаками, лошадьми, но маленькие зверьки, размерами меньше мышей, испытывают от него лишь незначительные неудобства. О насекомых и этого сказать нельзя: мухи и комары так легки, так мало весят, что способны плавать даже в воздухе, и тяготение их обременяет не больше, чем рыб»1.
Из этих высказываний следует, что невесомость — это такой фактор, к которому привыкли мухи и комары. Так почему же не может привыкнуть к ней Homo sapiens— человек разумный? К сожалению, все обстоит гораздо сложнее, чем об этом пишет Артур Кларк. Наши экспериментальные данные заставляют прийти к иному выводу, согласно которому длительная невесомость — это коварный враг, который может нанести удар тогда, когда этого никто не ожидает. Он будет действовать «исподтишка», но его удар вызовет катастрофические последствия. Ведь никто же не говорит, что можно привыкнуть к влиянию космической радиации. Нет оснований и для заключений о безобидности пребывания в состоянии невесомости.
Справедливость этих взглядов подтверждают данные, полученные во время полета американских космонавтов на космическом корабле «Джемини-7» и на побывавших в космосе подопытных собак Угольке и Ветерке. После 14-дневного пребывания в космосе у американских космонавтов было обнаружено повышенное выведение кальция из организма. {143}
Почему невесомость привела к нарушению минерального обмена? Почему началось усиленное выделение из организма именно кальция? Никто не мог дать объяснения этому явлению. Может быть усиленное выделение кальция из организма безразлично для организма? Нет, ибо это начало разрушения скелета, важнейшей составной частью которого является кальций. В результате нарушается прочность одной из основных структур организма, которая служит каркасом для организма, системой рычагов для передвижения, защитой таких нежных органов, как сердце, головной мозг.
Декальцинация скелета приводит к нарушению нормальной деятельности костномозговой части скелета, в которой созревают форменные элементы крови и синтезируется гемоглобин, следствием чего должно быть наступление анемии. Другими словами, оказывается уязвимым костный мозг, самая нежная эмбриональная ткань, сохраняющая свою эмбриональность в течение всей жизни организма,— ткань, запрятанная в самые глубокие недра организма. Костный мозг первым страдает при лучевом поражении. Следовательно, он является уязвимой структурой не только со стороны космической радиации, но и со стороны невесомости.
Уязвимость костного мозга со стороны космической радиации была известна давно, а об уязвимости со стороны невесомости пока даются только прогнозы.
Мы рассматриваем данные об усиленном выделении кальция из организма в условиях невесомости как первом доказательстве справедливости нашего прогноза, высказанного нами в 1963 г., то есть за несколько лет до полета «Джемини-7». Следует считать симптоматичным, что пребывание в состоянии невесомости даже в течение относительно короткого времени — 14 дней — уже создает весьма необычные условия для организма, нарушает все прежние механизмы адаптации к гравитационному полю. Отсутствие влияния сил гравитации раньше всего «замечает» скелет — система органов, испытывающая на себе в обычных условиях в максимальной степени действие этих сил.
Суть прогноза сводится к тому, что длительная невесомость — не безобидный фактор, к которому организм может привыкнуть. Этот прогноз пока единственный сигнал, что невесомость не безразлична для {144} организма, Что к неожиданным сюрпризам в Космосе со стороны невесомости надо готовиться на Земле. Сложность состоит, однако, в том, что искусственным путем пока невозможно воспроизвести условия длительной невесомости. Но именно поэтому этот, пока единственный прогноз, основанный на относительно небольшом фактическом материале, приобретает важное значение. Уверенность в его правильности подкрепляется еще и тем, что в земных условиях имеется ряд явлений, также получающих убедительное объяснение в свете изложенных выше взглядов.
Основной же вывод, который логически вытекает из сравнительного анализа имеющихся данных, сводится к тому, что тот замкнутый экологический комплекс, который должен быть создан в космическом корабле, предназначенном для длительных полетов на ближайшие планеты нашей солнечной системы, должен сохранять все основные элементы наших земных условий существования.
Наиболее острые дискуссии вызывает вопрос о составе атмосферы в отношении ее основного компонента — азота, составляющего почти 80% нашей атмосферы, который со времен Лавуазье считается инертным газом, безразличным для организма. На этом основании разрабатываются проекты замены азота более легким, но также инертным и безразличным гелием.
Даже если действительно азот был бы инертным и безразличным, то в этом случае утверждение о безвредности замены азота гелием весьма рискованно. Этот риск определялся бы тем бесспорным фактом, что вся эволюция органической жизни на нашей планете протекала в атмосфере азота. И совершенно неизвестно, какие коррелятивные связи в организме нарушились бы, если азот атмосферы был бы заменен гелием.
Однако открытие, сделанное профессором М. И. Волским в 1951 г. и официально зарегистрированное в 1968 г. в Комитете по делам изобретений и открытий при Совете Министров СССР, говорит о том, что азот атмосферы вовсе не безразличен для организма растений и животных, он усваивается организмами животных и растений в большей или меньшей степени. {145}
Следовательно, в свете этого открытия М. И. Волского вовсе не безразлично, будет ли в составе искусственной атмосферы космического корабля азот или его заменит гелий.
Примерно такое же положение создается и в случае длительной невесомости. То обстоятельство, что некоторые из животных организмов, такие как рыбы или мелкие насекомые, находятся на поверхности нашей планеты в состоянии, близком к состоянию невесомости, вовсе не дает права для заключения, что и другие организмы также в конце концов могут к нему привыкнуть. Все дело в том, что разные организмы по-разному решают проблему адаптации к невесомости.
Ведь первые выходцы на сушу среди позвоночных с огромными трудностями, путем колоссальной перестройки организма смогли, наконец, адаптироваться к наземным условиям, характерной особенностью которых является резкое усиление действия сил гравитации. Другие группы наземных позвоночных животных, филогенетически более молодые, смогли еще лучше адаптироваться к наземным условиям, изменив соответствующим образом структуру и функции организма.
Проблема освоения космоса выдвигает необходимость адаптироваться к жизни в условиях невесомости, с которой человек будет сталкиваться в межпланетных и межзвездных полетах. Как это будет осуществляться? Безусловно, не путем простого привыкания, ибо перестройка организма в процессе адаптации к гравитационному полю Земли зашла слишком далеко, чтобы можно было допустить такую возможность. По-видимому, пути адаптации человека к невесомости будут отличаться от адаптации животных, ибо они при этом меняют свою собственную природу. Человек же будет приспосабливаться с помощью техники, оставляя без изменений свою собственную природу.
Вывод таков, что невесомость — фактор весьма опасный, что пассивная адаптация к нему невозможна. Невольно вспоминается миф об Антее — сыне Посейдона и Геи (Земли), который побеждал в единоборстве всех своих противников, так как во время борьбы каждое прикосновение к Земле — его матери — придавало ему новые силы. Антей был побежден Гераклом, который поднял его на воздух, не дав ему прикоснуться к Земле. {146} Человечество, вступившее в эру освоения космоса, вновь сталкивается с проблемой, которая 20 с лишним веков назад нашла отражение в мифах древних греков. Теряя связь с Землей, с ее гравитационным полем, человек также будет терять силы, поскольку функционирование всех его структур приспособлено к гравитационному полю определенной интенсивности.
Одна из самых актуальных задач современной науки — изыскание способов, которые предотвратили бы вредные влияния длительного пребывания человека в состоянии невесомости.
| {147} |
Аксенова М. Я. Газообмен у северных оленей при голодании, кормлении и в период работы. Труды Бурят-Монгол. зоовет. ин-та, 1941, вып. 2.
Аршавский И. А. Физиология кровообращения во внутриутробном периоде. М.—Л., Медгиз, 1960.
Баркрофт Дж. Некоторые изменения, происходящие при рождении. «Успехи совр. биол.», 1935, т. 4, вып. 4—5.
Баркрофт Дж. Основные черты архитектуры физиологических функций. М., Биомедгиз, 1937.
Бернштейн А. Человек в условиях среднегорья. Алма-Ата, изд-во «Казахстан», 1967.
Боголюбский С. Н. О весовом росте скелета, мышц и внутренних органов в послеутробном онтогенезе «Советского мериноса» и других пород овец. Труды ИМЖ. 1951, вып. 35.
Бровар В. Я. Закономерности роста скелета домашних млекопитающих. Труды ТСХА, 1944, вып. 31.
Бровар В. Я. Силы тяжести и морфология животных. М., изд-во АН СССР, 1960.
Вернадский В. И. Биогеохимические очерки. М.— Л., изд-во АН СССР, 1940.
Вернадский В. И. Химическое строение биосферы Земли и ее окружения. М., «Наука», 1965.
Винберг Г. Г. Интенсивность обмена и пищевые потребности рыб. Изд-во Белгосуниверситета, 1956.
Винер Н. Творец и робот. М., изд-во «Прогресс», 1966.
Волский М. И. Новое в науке о дыхании. Горький, 1961.
Горб Т. Е., Семенченко 3. П. Повышение продуктивности романовских овец при длительном полноценном кормлении. «Советская зоотехния», 1950, вып. 2.
Дарвин Ч. Происхождение видов. М.— Л., Сельхозгиз, 1937.
Дранников С. Я. Некоторые наблюдения над газообменом у лягушек и других холоднокровных. Труды Харьковского об-ва испыт-ей природы, 1914, т. 47.
Дюрст И. Основы разведения крупного рогатого скота. М., Сельхозгиз, 1936.
Заварзин А. А. Очерки эволюционной гистологии крови и соединительной ткани, 1945, вып. 1.
Зенкевич Л. А. Очерки по эволюции двигательного аппарата {148} животных. Общие положения эволюции движения животных. «Журнал общей биологии», 1944, т. 5, вып. 3.
Ивлев В. С. Опыт оценки эволюционного значения уровней энергетического обмена. «Журнал общей биологии», 1959, 20, вып. 2.
Калабухов И. И. Спячка животных. Изд-во Харьков-го ун-та, 1956.
Кларк А. Черты будущего. М., изд-во «Мир», 1966.
Коржуев П. А. Дыхательная функция крови и скелет позвоночных животных. «Успехи совр. биол.», 1955, 39, вып. 2.
Коржуев П. А. Физиолого-биохимические аспекты невесомости. В сб.: «Медикобиологические аспекты невесомости». Изд-во «Медицина», 1968.
Коржуев П. А. Проблема невесомости с точки зрения земной физиологии. «Авиационная и космическая медицина». М., 1963.
Коржуев П. А. Гемоглобин. М., Изд-во АН СССР, 1964.
Коржуев П. А. Силы гравитации и филогенез позвоночных животных. «Успехи соврем. биологии», 1965, т. 60. вып. 2(5).
Коржуев П. А. Проблемы резервов кислорода в организме животных и человека. Сб.: «Кислородный режим организма и его регулирование». Киев, 1966.
Коржуев П. А. Гомеостаз и проблема невесомости, 1968, т. 13, № 4.
Коржуев П. А., Глазова Т. II. Количественная характеристика крови и кроветворных органов черноморских дельфинов. «Журнал эволюц. биохим. и физиол.», 1967, т 3, № 2.
Крепе Е. М. Особенности физиологии ныряющих животных. «Успехи соврем, биологии». 1941, т. 14, вып. 3.
Падучева А. Л. Особенности обмена веществ многоплодных маток. Вестник с.-х. науки, 1954, № 12.
Пшеничный П. Д., Дмитроченко А. П. Кормление сельскохозяйственных животных. Изд-во Сельхозлитература. М.—Л., 1961.
Ромер А. Ш. Палеонтология позвоночных животных. М.— Л., изд-во НТИ, 1939.
Рулье К. Ф. Общая зоология. Избр. биолог. произвед. М., изд-во АН СССР, 1954.
Северцов А. Н. Главные направления эволюционного процесса. Био-медгиз, 1934.
Северцов А. Н. Морфофизиологические закономерности эволюции. Изд-во АН СССР, 1939.
Сеченов И. М. Две заключительные лекции о значении так называемых растительных актов в животной жизни. «Мед. вестник», 1861, №26, №28.
Студитский А. Н. Потенциал скелетогенной клетки. Сборник, посвященный академику Насонову к 80-летию со дня рождения. М.—Л., Изд-во АН СССР, 1937.
Томме М. Ф. Обмен веществ и энергии у сельскохозяйственных животных. М., Сельхозгиз, 1949.
Хэммонд Р. Рост и развитие мясности у овец. М., Сельхозгиз, 1937.
Циолковский К. Э. Биология карликов и великанов. Сб.: «Путь к звездам», Изд-во АН СССР, 1961.
Чижевский А. Л. Аэроиниофикация в народном хозяйстве. М., Госпланиздат, 1960. {149}
Шмальгаузен И. И. Пути и закономерности эволюционного процесса. Изд-во АН СССР, 1940.
Шмальгаузен И. И. Проблемы дарвинизма. М.—Л., изд-во «Советская наука», 1946.
Acheson G. H., Dawes G. S. a. Mott J. С. Oxygen consumption and the arterial oxygen saturation in foetal and new-born lambs. J. Physiol., 1957, v. 135, N 3.
Allison A. C. Turnovers of erythrocytes and plasma proteins in mammals. Nature, 1960, v. 188, N 4744.
Brace K. Life span of the marmot erythroc. Blood, 1953, v. 8, N 7.
Brace K. a. Altland P. Red cell survival in the Turtle. Am. J. Physiol., 1955. v. 183. N 1.
Brace K. a. Altland P. Life span of the duck and chicken erythrocyte as determined with CH. Proc. Soc. Exp. Biol. Med., 195*5, v. 92 (3).
Bogert С. М. Thermoregulation in reptiles a factor in evolution. Evolution, 1949, v. 3.
Brock M. A. Production and life span of erythrocytes during hibernation in the golden hamster. Am. J. Physiol., 1960, v. 196 (6).
Brody S. Bioenergetics and growth with special reference to the efficiency complex in domestic animals. New York, 1945.
Cornelius C. E. a. Kaneko J. J. Erythrocyte survival studies in the mule deer, aoudad sheep, an springbok antelope, using glycine—2—Cu. Am. J. Veterin. Rev., 1945, v. 20 (78).
Cornelius C. E. a. Kamko I. J. Erythrocyte life span in the guanaco. Science, 1962, v. 137 (3531).
Delattre A. et Fenart R. L'evolution du crane et le cosmos. Cahiers des etudes biol., 1960. N 6-7.
Foxon G. Problems of the double circulation in vertebrates. Biol. Rev., 1955, v. 30 (2).
Gray I. Studies in the mechanics of the tetrapod skeleton. J. Exp. Biol., 1944, v. 20(2).
Gregory W. Grandfather fish and his descendants. Natural History, 1941, v. 48.
Groebbels F. Der Fogel. Bau, Funktion Lebenserscheinung Einpassung. 1937, v. 1.
lleberich F. The functional separation of venous and arterial blood in the univ'entricular frog heart. Ann. New York Acad. Sci, 1965, v. 127, N 3.
Hesse R. Das Herzegewicht der Wibeltiere. Zool. I. Abt. Allg. Zool. u. Physiol, d. Tiere, 1921, v. 38.
Irving L. Respiration in diving mammal;. Physiol. Rev., 1939, v. 19.
Jordan H. Comparative haeinatology. Downey's handbook of Haematology, 1938, v. 2.
Krogh A. On the cutancow and pulmonary respiration of the frog. Skand. Arch. Physiol., 1904, v. 15.
Lawrie R. The activity of the cytochrome in muscle and its relation to myoglobin. Biocli. J., 1933, v. 55, 298.
Macallum A. B. The Paleochemistry of the body fluids and tissues. Physiol. Fev., 1.926, v. 6.
Mechanick N. Untersüchungen über das Gewidit des Knochenmarkes der Menschen. Ztschr. Ges. Arat., 1926, B. 79. {150}
Letnez L. a. Kopecky H. the number of erythrocyte recirculation in chick embryo and hen, a note on embryonic heart efficiency. Physiol. Bohemoskov., 1962, v. 11, N 2.
Robinson D. The muscle hemoglobin of seals as an oxygen store in diving. Science, 1939, v. 90.
Scholander P. F. Experimental investigation on the respiratory function in diving mammals and birds Hvalradets Skrifter, 1940, N 22.
Szarski H. Pochodzenie plazow. Warszawa, 1961.
Szarsky H. The structure of respiratory organs in relation to Body Size in Amphibia. Evolution, 1964, 18, N 1.
Thompson D. Growth and form. Cambridge Univ. Press, 1917.
Varicak T. Medulla ossium Urodela. Biol. Hrvaltsko priradasl drustvo, 1958, v. 11.
Wintrobe M. Variations in the size and hemoglobin content of erythrocytes in the blood of various Vertebrates. Folia Haematologica, 1933, v. 51.
| {151} |
1 В. И. Григорьев и Г. Я. Мякишев. Силы в природе. «Наука», М., 1966, стр. 52.
1 «По подсчету Осборна, который берет по всей вероятности слишком низкие цифры, мезозойская эра продолжалась около 9 млн. лет; третичная — около 3 млн. лет, т. е. втрое короче. По современным данным (1934), как указал мне В. И. Вернадский, эти цифры по всей вероятности должны быть гораздо более значительными, а именно 220 млн. лет для мезозоя и 80 млн. лет для третичного периода».
2 А. Н. Северцов. Главные направления эволюционного процесса. Биомедгиз. М., 1934, стр. 122.
1 И. М. Сеченов, О так называемых растительных актах в животной жизни. «Медицинский вестник», 4861, № 26, стр. 235; № 28, стр. 253.
1 В. И. Вернадский. Химическое строение биосферы Земли и ее окружения, М., «Наука», 1965, стр. 36.
2 Там же, стр. 37.
1 В. И. Вернадский. Биогеохимические очерки. М.—Л., Изд-во АН СССР, 1940, стр. 19.
2 Там же, стр. 127.
1 В. И. Вернадский. Биогеохимические очерки. М.—Л., Изд-во АН СССР. 1040, стр. 139.
2 И. М. Сеченов. О так называемых растительных актах в животной жизни. «Медицинский вестник», 1861, № 26, стр. 235: № 28, стр. 233.
1 К. Э. Циолковский. Биология карликов и великанов. Сб.: «Путь к звездам». М., Изд-во АН СССР. 1961, стр. 311.
1 В. Я. Бровар. Силы тяжести и морфология животных. М., Изд-во АН СССР, 1960, стр. 239.
1 Н. И. Калабухов. Спячка животных. Изд-во Харьковск. ун-та. 1956.
1 П. Л. Коржуев. Происхождение гомойотермии. «Успехи соврем, биологин», 1940, т. 12, № 3.
1 В. Я. Бровар, Закономерности роста скелета домашних млекопитающих. Труды ТСХА, 4944, вып. 31, стр. 185—204.
1 В. И. Вернадский. Биогеохимические очерки. М—Л.. Изд-во АН СССР, 1940, стр. 124.
1 П. А, Коржуев. Дыхательная функция крови и скелет позвоночных животных. «Успехи соврем, биологии», 1955, т. 39, стр. 163—182.
1 Д. Хеммонд. Рост и развитие мясности у овец. М., Сельхозгиз, 1937, стр. 372.
3 И. Дюрст. Основы разведения крупного рогатого скота. М., Сельхозгиз, 1936. стр. 455.
А. Ш. Ромер. Палеонтология позвоночных животных. М.—Л., ГОНТИ, 1939, стр. 169.
И. И. Шмальгаузен. Пути и закономерности эволюционного процесса. Изд-во АН СССР. 1940. М.—Л., стр. 165.
1 И. И. Шмальгаузен. Проблемы дарвинизма. М., «Сов. наука», 1946, стр. 528.
2 Ч. Дарвин Происхождение видов. М.—Л., Сельхозгиз, 1937. стр. 276.
А. П. Виноградов. Введение в геохимию океана. М., изд-во «Наука», 1967, стр. 213.
1 Д. И. Северцов. Главные направления эволюционного процесса. М, Биомедгиз. 1934, стр. 150.
2 И. И. Шмальгаузен. Пути и закономерности эволюционного процесса. М„ Изд-во АН СССР, 1940, стр. 230.
1 Д. Н. Северцов. Главные направления эволюционного процесса. М.—Л., Биомедгиз, 1934, стр. 150.
Д. Н. Северцов. Главные направления эволюционного процесса. М.—Л., Биомедгиз, 1934, стр. 72.
Л. Л. Зенкевич. Очерки по эволюции двигательного аппарата животных, «Журнал общей биологии», 1944, т. 5, № 3, стр. 129.
1 В. Б. Суханов. Эволюция движений позвоночных животных. «Наука и жизнь», 1968, № 7
1 П. Л. Коржуев. Проблема резервов кислорода в организме животных и человека. В сб. «Кислородный режим организма», Киев, 1966.
1 Б. В. Нейман. Расширяющаяся Земля, М., 1962.
2 Л. С. Смирнов, Ю. Н. Любина, О возможности изучения изменения силы тяжести с геологическим временем. ДАН СССР, т. 187, вып. 4.
1 Норберт Винер — основатель кибернетики придерживался другой точки зрения. Он писал (1966 г.): «Силы земного притяжения столь же дружественны нам сколь и враждебны».
1 А. Кларк. Черты будущего. М., «Мир», 1966, стр. 287.
3 | |
5 | |
Глава I | 9 |
Глава II | 41 |
Глава III | 64 |
Глава IV | 105 |
Освоение водной среды — переход в «гипогравита- | |
Глава V | 124 |
Жизнь без гравитации — проблема невесомости — |
|
Петр Андреевич Коржуев Эволюция, гравитация, невесомость Утверждено к печати редколлегией серии научно-популярных изданий Академии наук СССР Редактор издательства В. Н. Вяземцева. Художник В. Ф. Соболев Художественный редактор В. Н. Тикунов Технический редактор В. В. Волкова Сдано в набор 9/II 1971 г. Подписано к печати 2/VI 1971 г. Формат 81×1081/32. Бумага № 1. Усл. печ. л. 7,98. Уч.-изд. л. 7,6. Тираж 12000 экз. Т-09242. Тип. зак. 4487. Цела 49 к. Издательство «Наука». Москва К-62, Подсосенский пер., д. 21 2-я типография издательства «Наука». Москва Г-99, Шубинский пер., 10 |
