
Т.Г.МОРГАН
РАЗВИТИЕ
и
НАСЛЕДСТВЕННОСТЬ
Перевод Ю. Я. КЕРКИС
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО
БИОЛОГИЧЕСКОЙ И МЕДИЦИНСКОЙ ЛИТЕРАТУРЫ
МОСКВА—ЛЕНИНГРАД
1937
| << | {1} | >> |
57 М 79
EMBRYOLOGY AND GENETICS
BY
Т. Н. MORGAN
1934
Книга Моргана, крупнейшего американского биолога, излагает популярным языком труднейшие, далекие еще от разрешения, но оживленно разрабатываемые вопросы значения наследственности в развитии. Книга рассчитана на широкие круги читателей, желающих ознакомиться с теоретическими основами одной из наиболее актуальных областей современной биологии — учением о развитии, в связи с современными представлениями о наследственности.
| << | {2} | >> |
Автор книги, крупнейший современный генетик, Т. Г. Морган еще в начале текущего столетия с большим мастерством и энергией, вместе с Ж. Лебом, Вильсоном и другими, выступил горячим защитником экспериментального метода, начавшего тогда широко внедряться в практику биологического исследования. В противовес господствовавшим в то время сравнительным и описательным исследованиям, создавалось новое направление — «экспериментальная зоология», адепты которого противопоставляли, по их мнению, «туманным» биологическим теориям «точные» данные эксперимента. Большой заслугой Моргана и его единомышленников является то, что они повели решительную борьбу с попытками виталистов декларировать принципиальную невозможность решения естественно-научными методами основных биологических проблем. Находясь на материалистическом пути, Морган, однако, все же не выходил из рамок эмпирического механицизма. Морган полагает, что материалистическое исследование биологических явлений, в конечном счете, должно привести к сведению их к законам физики и химии. Как это характерно для большинства представителей «механистического» объяснения в науке, Морган пренебрегает или отодвигает на второй план всякие чисто биологические закономерности, особенно исторические, полагая, что невозможность формулирования их в понятиях физики и химии свидетельствует об их слабости и незавершенности, которые предстоит еще преодолеть. Поэтому, когда Морган касается таких особенностей организма, как, например, целесообразность, столь характерная для книги в целом ясность изложения теряется. Так, трудно понять в точности, что именно хотел высказать Морган, утверждая, что «отбор является одним из наиболее неясных явлений, свойственных живым существам» (стр. 12). Не менее странно звучит и утверждение о «запутанности» биогенетического закона, который, по мнению Моргана, {3} «скорее служит для сокрытия истинного родства» (стр. 153). Однако полемические высказывания Моргана о биогенетическом законе бьют мимо цели, так как этот закон отнюдь не гласит, как это утверждает Морган, что «эмбрионы представителей высших групп проходят через взрослые стадии более низко организованных представителей этой же группы» (стр. 153). В современном понимании этого закона не содержится требования о повторении взрослых стадий, речь идет о наличии при эмбриональном развитии зачатков ряда признаков, соответствующих строению предков.
Повышенные требования, предъявляемые у нас к обсуждению биологических проблем, связанных с человеком, остаются вне поля зрения Моргана. Неудивительно поэтому, что для советского читателя будет неприемлемо в частности утверждение Моргана, что «усовершенствование тестов психологического поведения... позволит нам лучше различать прирожденные способности от результатов внешнего влияния общественных отношений и воспитания» (стр. 115). Понятно, что успешность подобных исследований меньше всего зависит от порочного в своей основе метода тестов.
Отдельные неверные утверждения с избытком искупаются в высокой степени оригинальным изложением, смелыми и часто неожиданными сопоставлениями казалось бы разнородных явлений, умелым использованием разнообразнейшего биологического материала и общим высоким уровнем изложения. Несомненно, что книжка Моргана, излагающая доступным языком наиболее трудные проблемы современной биологии, по праву займет одно из первых мест в популярной биологической литературе и окажется полезной и советскому читателю.
| << | {4} | >> |
Мне всегда казалось странным, что в то время как вопрос об эволюции возбуждает всеобщий интерес, развитие индивидуума из яйца вызывает так мало любопытства. Быть может, чудесное происхождение человека и тайны, окружающие его появление на земле, составляющие обычный элемент фольклора многих народов, достаточно объясняют интерес к этому людей, питавшихся подобными рассказами с детства. Между тем прозаический факт, что цыпленок развивается из куриного яйца, принимается скорее как банальность, а не как интереснейшая проблема.
Если происхождение на земле жизни, кульминационной точкой эволюции которой явилось образование человеческих рас, на первый взгляд требует чуда, то такое же или, может быть, несколько иное чудо требуется, повидимому, всякий раз при развитии яйца во взрослый организм. Если бы мы могли представить наблюдателя с ограниченными человеческими способностями, присутствовавшего в течение тех миллионов лет, когда совершалась эволюция растений и животных, то сомнительно, был ли бы он лучшим свидетелем происходящего вокруг него, чем современный палеонтолог. В горных породах на поверхности земли действительно сохранились фрагменты эволюции высших групп. Кости и раковины представителей этих групп, расположенные в их вероятной последовательности, рисуют картину того, что происходило. Правда, сохранилась только небольшая часть того, что было, но все же эта часть достаточна для того, чтобы послужить ключом к установлению последовательности событий. Но попытки палеонтологов найти причинные факторы, вызвавшие эти изменения, в большинстве случаев приводили к непоследовательным заключениям, немногим более убедительным, чем попытки народностей, стоящих на низкой ступени своего развития, объяснить физику солнечных затмений или физиологическое действие лекарственных трав. {5}
Эмбриолог, наблюдающий изменения формы в течение эмбрионального развития яйца, в результате которых организм приобретает сходство со своими родителями, находится в немного лучшем положении, чем воображаемый наблюдатель, присутствовавший в течение эволюции организмов, живущих в наши дни; ибо, как и последнему, ему не видно почти ничего из того, что лежит в основе наблюдаемых изменений. Умозрительные построения такого эмбриолога не смогли уложить явления развития в рамки точной науки. Лишь начав применять методы, с помощью которых наука отделяет пшеницу от плевел, т. е. начав применять рабочие гипотезы, проверяемые количественными исследованиями, или, одним словом, экспериментальные методы, эмбриология стала превращаться в точную науку. Влечение современного эмбриолога к экспериментальному методу связано не столько со стремлением открыть, насколько химия и физика способны объяснить ему, что лежит в основании явления, как стремлением использовать экспериментальный метод в качестве орудия для вскрытия природы видимых явлений развития. Если на последующих страницах уделено достаточно внимания чисто внешней стороне развития, то это сделано только постольку, поскольку это необходимо для того, чтобы выяснить сущность тех опытов, которые должны поднять проблему на более высокую ступень.
После того как в 1900 году стало известным открытие Менделя, наступид один из наиболее замечательных периодов развития биологии во всей ее истории. В настоящее время основные законы наследственности уже известны и так как наследственные свойства особи передаются из поколения в поколение именно через яйцо, то важность понимания явлений развития для дополнения знания законов наследственности очевидна, и связь этих двух ветвей экспериментальной биологии стала предметом углубленного интереса. На нижеследующих страницах я попытался в популярной форме установить их взаимоотношения. Слишком очевидно, что остается сделать еще очень многое, но открытия, полученные при помощи экспериментального исследования наследственности и эмбриологии, говорят за то, что мы можем сделать значительно больше уже достигнутого.
Пасадина, Калифорния, 2 февраля 1934 г.
Томас Гент Морган
| << | {6} | >> |
Осадочные породы земной поверхности вскрывают самую последнюю часть длинного эволюционного пути ныне живущих животных и растений. В то время как потребовались миллионы лет для того, чтобы эти изменения могли совершиться, развитие каждого индивидуума из сравнительно просто организованного яйца во взрослый организм, обнаруживающий внешне видимое сложное строение, является делом только дней или даже часов. Однако это сравнение может быть ошибочным, так как в процессе эволюции, вероятно, были длинные периоды, в течение которых у видов не происходило либо никаких изменений, либо только небольшие изменения. Каждый же дальнейший шаг у отдельных особей мог в действительности происходить в бесконечно малый отрезок времени, от гена к гену, путем внезапных изменений в одной из единиц наследственности.
Отождествление яйцевых клеток с одноклеточными предками, из которых эволюционировали высшие формы, требует обоснования. Возможно, что ближе к истине обратное положение, а именно, что яйцо ныне живущего организма настолько отлично от исходного одноклеточного предка, насколько современный взрослый организм отличается от своего взрослого предка. Оба эти положения требуют оговорок, ибо все сводится к тому, что подразумевается под сходством и различием. В яйце имеются все возможности для быстрого развития признаков взрослой формы и в этом смысле яйцо неизмеримо отлично от исходного одноклеточного предка. Эти отличия сводятся к различию в наследственных единицах: простейшее и яйцо несколько похожи друг на друга только своими видимыми особенностями. Так как мы ничего не знаем о различиях в строении между наследственными элементами исходного {7} простейшего и яйца ныне живущего организма, то нелепо пытаться производить какое-либо серьезное сравнение относительной сложности обоих. Они обнаруживают только поверхностное сходство своих внешне видимых структур.
Однако мы можем произвести сравнение в другой плоскости. Прародительский тип должен был претерпевать меньшее число видимых изменений при развитии от яйца до взрослого организма, ибо у размножающихся делением одноклеточных форм — протистов — каждой дочерней клетке практически достаточно лишь увеличиться до исходных размеров, чтобы стать подобной материнской, а у низших многоклеточных организмов число стадий после деления яйца очень мало по сравнению с таковым у высших форм. Но даже в этом случае сравнение может быть ошибочным, ибо у высших форм принимаются в соображение именно видимые изменения, которые мы наиболее часто представляем себе как изменения формы или структуры, тогда как физиологические процессы у одноклеточных и многоклеточных типов являются, вероятно, много более сходными. У высших форм эти процессы разделены по системам органов, но они могут быть почти такими же, как и у простейших. Описательная эмбриология занимается исключительно изменениями формы и очень мало физиологией развития. Несмотря на то, что всегда некоторые ученые интересовались физиологией зародыша позвоночных, особенно на поздних стадиях его развития, только недавно последнему стало уделяться серьезное внимание.
Много лет назад, примерно между 1850 и 1900 годами, эмбриологи были увлечены идеей, что при развитии высших форм полностью повторяется исторический путь, по которому протекала их эволюция. Эта идея приобрела известность под именем теории рекапитуляции. Под влиянием этой теории была проведена огромная чисто описательная эмбриологическая работа и в настоящее время известна эмбриология всех типов животных, часто до самых мелких деталей. В маленьких прозрачных яйцах стадии развития могут быть прослежены под микроскопом, но даже и для менее прозрачных яиц были придуманы технические приемы, которые позволили наблюдать изменения, происходящие под поверхностью яйца. Совершенствование этих методов — окраска, заключение в парафин, производство тонких срезов, заделывание их в бальзам на предметные стекла и реконструирование {8} целого эмбриона из воска — долгое время занимало внимание большого числа профессиональных эмбриологов, что отвлекало их от рассмотрения физических и химических явлений, лежащих в основе наблюдаемых стадий развития. Влечение к истории было непреодолимо, в особенности для того, кто считал, что все видимое и описываемое им является историей «творения», или, как это называлось, эволюцией. Вскоре накопилось очень много данных, касающихся развития всех главных форм животных, для чего потребовалось столь же тщательное наблюдение, как и во всех изобразительных искусствах. В многочисленных монографиях появились прекрасные рисунки, иллюстрирующие развитие зародыша из яйца. Чем лучше был художник, тем более блестяще было их оформление. Анатомия развития стала так же хорошо изученной, как и более старая анатомия взрослых структур, для изучения которых точно так же требовалось непосредственное наблюдение и искусство делать цветные и перспективные иллюстрации.
В течение последних лет прошлого столетия и вплоть до настоящего времени развивалась новая отрасль, называемая экспериментальной эмбриологией, или, иногда, механикой развития. Частично ответственной за это изменение интереса была реакция против старой интерпретации стадий развития как рекапитуляции предков. Новые способы исследования процессов, происходящих за кулисами, открытие никогда ранее не предполагавшихся потенций яйца, применение различных агентов для того, чтобы вызвать искусственные изменения в развитии, выяснение роли внешней среды в нормальном развитии — все это способствовало пробуждению новых интересов биологии.
Много молодых эмбриологов с энтузиазмом занялись этой новой областью исследования. Очень многое было выяснено и возникло очень много проблем, весьма отличных по своему характеру от тех, которые занимали умы предыдущего поколения ученых. Казалось, что здесь была возможность дальнейшего движения к познанию процессов развития, и мысль, что эмбриология может быть поставлена на экспериментальную основу, была особенно притягательна для тех, кто был знаком с большими успехами, вызванными экспериментальным методом в химии и физике. Эмбриологу казалось, что он имеет дело с настолько отличными проблемами, что не представлялось {9} возможным сразу применить к ним законы химии и физики. Он имел дело с такими сложными вещами и вопросами, как белки, коллоиды, силы, действующие на поверхностях, проницаемость и т. д., которые сами физики еще не привели в соответствие с другими областями их работы. Действительно, почти все так называемые экспериментальные работы в эмбриологии оставались еще на биологическом уровне. Стали известными многие моменты в развитии яйца, которые до этого никогда не предполагались, но привлечение физики и химии со стороны так называемых механиков развития чаще было основано на аналогии, чем на демонстрации, и даже «химическая эмбриология» была в значительной степени просто описанием различных химических соединений, найденных в яйце и зародыше. Правда, превращение некоторых из этих соединений в другие вещества, или в конечный продукт, является существенной частью эмбриологических пробл м, но эмбриолог столь же много занимается типом реакций, приводящих к определенным изменениям в форме зародыша, как и происхождением одних веществ из других.
Замечательный факт, что яйцо, внешне с очень несложной видимой организацией, развивается в сложную взрослую форму, обладающую огромным числом структур, привлекал интерес философов от Аристотеля до Уайтхэда, которые в общей форме ясно представляли себе, что здесь происходит нечто таинственное, не имеющее себе параллели в других областях научного знания. Эти мыслители находились главным образом под впечатлением тех сторон организации, которые выражаются в форме, как наиболее важной особенности развития, и даже в наши дни эти изменения формы остаются самой выдающейся особенностью развития. При этом возможность зависимости этих изменений формы от химических изменений, происходящих в зародыше, либо принималась на веру, либо же вовсе игнорировалась.
Более всего дискутировавшийся философский «принцип» известен под именем энтелехии. Энтелехия, являющаяся, повидимому, тем же понятием, которое фигурировало под этим названием в учении Аристотеля, постулировалась как принцип, ведущий развитие к определенной цели, как нечто внешнее и не зависящее от химических и физических свойств веществ яйца, как нечто такое, что, не влияя на изменения энергии, направляет или регулирует такие {10} изменения почти так же, как человеческий разум может контролировать ход или постройку машины. Принятие такого принципа сделало бы бесполезным применение экспериментального метода для изучения развития, так как оно направлялось бы и контролировалось энтелехией. Более новое учение об «организме как целом» по существу не очень сильно отличается от учения об энтелехии, поскольку мы не можем представить себе другие пути ультра- или сверхматериалистической координации целого.
Поэтому, если не принять, что законы развития являются отличными от законов физики, в самом широком и наиболее современном употреблении этого выражения, то, казалось бы, лучше оставить эти метафизические вопросы и попытаться, несмотря на все потребное для этого количество времени и труда, выяснить, насколько знание химических и физических изменений, происходящих в яйце, сможет приблизить нас к пониманию процессов развития. Может оказаться, конечно, что для понимания находящейся в яйце системы, до сих пор называемой иногда организацией яйца, потребуются относительно новые законы, свойственные системам коллоидов, эквилибрированным солевым растворам, полупроницаемым оболочкам, границам фаз и т. п. Но если к этим «принципам» все же приложимы законы физики, и химии, будь то старые или новые, то эмбриология окажется в одном ряду с другими естественными процессами, понимаемыми в широком смысле этого слова, касающимися как живого, так и неживого вещества. Если же выяснится, что для познания живого вещества требуется нечто для физических наук совершенно новое, то будет своевременным выяснить вопрос о естественности или неестественности этого нечто. Тем временем же, повидимому, ясно, что ближайшим шагом должна быть настойчивая попытка познать все, что возможно, о свойствах системы или структуры, составляющей яйцо. Это не значит, что мы должны полностью прибегнуть к тем же способам анализа, которые были выработаны физиками и химиками для изучения веществ, с которыми они имеют дело, а лишь только то, что мы не должны отвергать ни одной возможности глубже проникнуть в поведение таких систем с помощью биологического эксперимента.
До тех пор нельзя с уверенностью утверждать, что физико-химические проблемы отличаются от биологических, пока мы не узнаем больше о последних. Для каждого {11} изучающего эмбриологию должно быть очевидным, что мы только начали получать сведения о, так сказать, биологическом «строении» яйца и что до сих пор мы еще очень мало знаем о химии и физике развития. Если бы оказалось, что ни классическая механика, ни новейшая физика недостаточны для объяснения явлений развития, то будет по крайней мере подготовлена почва для открытия каких-то новых законов, применимых к живой материи. Но до тех пор, пока не будет показано, что то, что мы называем свойством или свойствами живой материи, не имеет совершенно ничего общего с тем, что называется неживыми системами, будет, пожалуй, близорукостью пользоваться ходячими метафизическими принципами, или принимать их хотя бы временно. Именно эта альтернатива разделяет тех, кого по настоянию философов называют механистами, и тех, кого биологи называют метафизиками. Нет нужды пытаться найти компромисс, говоря, что каждый имеет свою собственную область. Мистицизм с точки зрения ученого в настоящее время является совершенно устарелым путем решения изучаемых им проблем.
Однако большинство современных биологов не столько находятся под впечатлением идеи существования особого закона жизни, как увлечены большим разнообразием явлений, обнаруживаемых, живой материей. Им кажется столь же преждевременным, как и претенциозным обсуждать какие-то воображаемые идеальные свойства жизни, в то время, когда имеется огромное число фактов, говорящих о чрезвычайном разнообразии свойств живой материи, причем каждое из этих свойств настойчиво требует своего объяснения до каких-либо попыток синтезировать из них жизнь. Можно, конечно, выбрать одно или несколько таких свойств, например, разум, целесообразность или свободную волю, и сделать их sine qua поп живой материи, но нельзя пройти и мимо того факта, что отбор является обычно одним из наиболее неясных явлений, свойственных живым существам. Формообразующие силы, полярность, симметрия и целесообразная регуляция являются примерами подобных свойств, с которыми мы встречаемся в эмбриологии.
Генетика и экспериментальная эмбриология стали настолько тесно переплетаться друг с другом, что обе науки могут быть в настоящее время до некоторой степени объединены. Правда, еще желательно достигнуть их соприкосновения {12} во многих важных точках, но уже достаточно известно для того, чтобы попытаться эмбриологию и генетику излагать одновременно. Хотя каждая отрасль развивалась в значительной части независимо от другой, тем не менее их взаимозависимость в настоящее время настолько очевидна, что генетик принимает на веру основные факты эмбриологии, а эмбриологу приходится признать свою зависимость от данных, доставляемых генетикой. Так, например, деление клетки и поведение хромосом во время созревания яиц и спермиев послужило рабочей схемой для теории наследственности. Сведения об изменениях, происходящих во время созревания яиц и спермиев, получены эмбриологией. Обратно, генетический анализ позволил нам проникнуть глубже этих видимых изменений, — в строение самих хромосом. Общая точка соприкосновения эмбриологии и генетики заключается во взаимоотношении между наследственными единицами в хромосомах, т. е. генами, с протоплазмой клетки, в которой влияние генов приобретает свое видимое выражение. Говоря о способе функционирования генов во время развития, я в дальнейшем изложении везде, где это только возможно, оттеняю две крайних точки зрения и указываю третью. Допущение, которое делается в большинстве генетических интерпретаций, заключается в том, что все время одинаково действуют все гены. Однако, если развитие зависит только от генов, то это допущение не объясняло бы, почему одни клетки зародыша развиваются одним путем, а другие другим. Противоположная точка зрения должна была бы предполагать, что по мере протекания развития приходят в действие различные группы генов. Первая точка зрения, что все гены все время действуют одинаково, оставляет проблемы эмбриологии на той же стадии, на которой они, повидимому, всегда и находились, т. е. сводит все к протоплазме. Противоположный взгляд мог бы, пожалуй, дать формальное объяснение развития, но он не находится в соответствии с результатами, полученными при изменении последовательных плоскостей дробления путем сдавливания. Ру и Вейсман пытались объяснить развитие довольно сходным образом, допуская, что заключающиеся в хромосомах детерминанты качественно распределяются во время развития. Однако и в то время не было данных в пользу этого взгляда, а в настоящее время многое противоречит ему. Мысль, что различные наборы {13} генов приходят в действие в различное время, может быть подвергнута серьезной критике, если не смогут быть приведены некоторые основания в пользу этого заключения: Следующее замечание снимает эти возражения. Известно, что протоплазма в различных частях яйца несколько различна и что по мере дробления эти различия становятся более очевидными, благодаря происходящему при этом перемещению веществ. Из протоплазмы образуются вещества, необходимые для роста хроматина, и из ее же веществ гены вырабатывают свои продукты. Можно предполагать, что исходные различия в протоплазматических участках влияют на активность генов. В свою очередь гены будут затем влиять на протоплазму, в которой начнется новый ряд взаимных реакций. Таким образом, можно себе представить постепенное усложнение и диференцировку различных участков зародыша.
| << | {14} | >> |
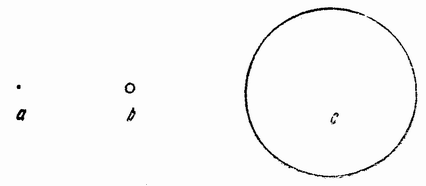
Еще задолго до существования какой-либо летописной истории человечества должно было быть известно, что некоторые животные развиваются из яиц, — по крайней мере те животные, яйца которых видны невооруженным глазом. Птицы, ящерицы, лягушки и рыбы имеют крупные яйца; у большинства других животных яйца мельче, причем у многих они еле видны невооруженным глазом (рис. 1). Размеры человеческого яйца составляют лишь одну пятую миллиметра, и так как яйцо развивается внутри материнского организма и, следовательно, скрыто, то происхождение человеческого эмбриона оставалось до тех пор тайной, пока Бэр в 1820 году не установил существование у млекопитающих яиц.
Значительно более трудным было установить роль самца в зачатии. Необходимость семени для начала развития яйца могла подозреваться еще в очень давние времена, но о самой существенной части семени не могло создаться никакого представления до тех пор, пока в 1677 году Гам и Левенгук не открыли существование в семени анималькулей или сперматозоидов. Однако еще в течение долгого времени не являлось общепризнанным, что эти микроскопические анималькули являются действительными агентами в развитии. Только спустя 177 лет (в 1854 г.) Ньюпортом было обнаружено проникновение спермия в яйцо и с этого времени стало признанным, что яйцо и сперматозоид являются клетками.
Следующий шаг был сделан в 1875 г. Гертвигом, который обнаружил, что головка спермия после проникновения в яйцо увеличивается, становясь хорошо различимым ядром, которое сливается с ядром яйца. Таким образом, два соединившихся ядра дают начало ядру делящегося яйца, которое является предшественником всех ядер в клетках образующегося {15} эмбриона. Так как предполагалось, что ребенок наследует поровну от обоих родителей и так как сперматозоид привносит с собой только ядро,— его хвостик, как правило, в яйцо не проникает — то из этого был сделан вывод, что наследственные вещества содержатся в ядре. В период, к которому относится этот вывод, было уже известно, что всякий раз когда ядро подготовляется к делению клетки, в нем возникают палочкообразные или V-образные тельца, имеющие сильное сродство к некоторым краскам и потому названные хромосомами. Величина,
 |
|
Рис. 1. Диаграмма относительной величины яиц ланцетника (а), лягушки (b) и желтка куриного яйца (с). |
число и форма хромосом характерны для каждого вида. Способ, при помощи которого поддерживается это постоянство, стал понятен, когда Ван Бенеден (1883) сделал открытие, что ядро спермия привносит в яйцо половину типичного числа хромосом — отцовские хромосомы, и что ядро яйца перед оплодотворением тоже содержит в себе половинное число хромосом — материнские хромосомы. Другими словами, каждое зрелое яйцо и сперматозоид содержат ровно половину того числа хромосом, которое характерно для данного вида. Каким образом происходит редукция числа хромосом в зрелых половых клетках, было открыто позднее, в результате изучения изменений, имеющих место в яйцах и семенных клетках на последних стадиях их развития. Эти изменения будут описаны в следующей главе.
Анализ передачи по наследству родительских признаков показал, что хромосомы не являются конечными единицами наследственности, а содержат в себе более мелкие единицы, называемые генами. Последние расположены в {16} каждой хромосоме в линейном порядке, подобно бусам, нанизанным на нитку. Данные, что гены являются конечными наследственными единицами, покоятся не на прямом наблюдении, так как гены находятся за пределами видимости наших микроскопов, а на выводах из фактов наследственности.
К этому краткому обзору основных ступеней, приведших к современной концепции механизма наследственности, должно быть присовокуплено также и другое направление исследований, которое помогло выяснить роль хромосом в наследственности. Данные эти исходят из экспериментальной эмбриологии. Если яйцо разрезано перед оплодотворением на две части, то каждая из них может быть оплодотворена одним сперматозоидом и может развиться в зародыш. Одна часть яйца содержит ядро и ее развитие происходит так же, как и развитие целого яйца; она содержит после оплодотворения полное, диплоидное число хромосом. Другая же часть, вначале не имевшая ядра, после проникновения в нее спермия получает ядро сперматозоида с половинным числом хромосом, по одной каждого сорта. Это число хромосом является гаплоидным. И эта часть так же развивается и ее развитие устанавливает тот факт, что один набор хромосом достаточен для осуществления нормального развития. Правда, такие «гаплоидные» эмбрионы оказываются иногда слабыми. По крайней мерз в некоторых случаях было показано, что эмбрионы, начавшие свое развитие с половинным числом хромосом, достигнув взрослых стадий, удвоили число хромосом путем задержки одного из первых делений протоплазмы яйца. Тем не менее тот факт, что из гаплоидного куска может тоже развиться типичный эмбрион, показывает, что одного набора достаточно для образования характерных черт особи. В подтверждение этого вывода имеются также данные, полученные при изучении партеногенетического развития.
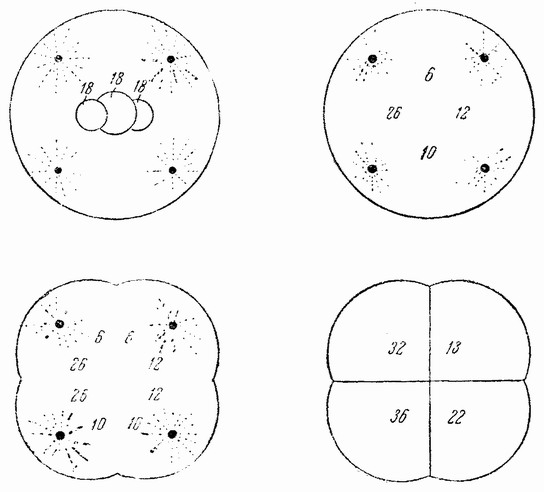
Другое доказательство значения хромосом было получено остроумным опытом Бовери. Было известно, что при проникновении в яйцо морского ежа двух сперматозоидов, яйцо при первом делении делится на три или четыре клетки вместо двух, как это происходит при нормальном развитии. Такие яйца не развиваются нормально. Как показал ранее Гертвиг, в таких диспермических яйцах первое деление хромосом происходит неправильно, и возникающие три или четыре клетки получают различные числа хромосом. {17} В то время было известно из результатов некоторых опытов Дриша, что если первые две клетки яйца морского ежа отделить друг от друга, то из каждой образуется нормальный эмбрион. То же самое имеет место при разделении четырех клеток. В этих случаях каждая клетка содержит, конечно, обычный двойной набор хромосом. Бовери же в противоположность этому нашел, что при разделении трех клеток диспермического яйца только одна из них, и то редко, развивается в нормальный зародыш (рис. 2). Он пришел к заключению, что неспособность большинства таких изолированных клеток нормально развиваться обусловлена неправильным распределением хромосом и что нормальный эмбрион развивается только из тех клеток, которые случайно получили по крайней мере один полный набор хромосом. Это заключение верно только отчасти. Одного набора хромосом действительно достаточно для развития, но если сверх него присутствуют также и другие хромосомы, то развитие оказывается нарушенным. В условиях опыта только редко может случиться, что в одну из трех клеток попадает в точности один набор хромосом и ничего больше. Так как превышение одного набора на любое число хромосом является нарушающим фактором, то для того, чтобы сделать достоверными цифровые отношения, на которых Бовери основывает свои выводы, необходим более подробный анализ способа распределения хромосом в таких яйцах. Позднее, генетическими опытами было показано, что основной набор может быть увеличен в три, четыре или большее число раз и тем не менее давать нормальное развитие, в то время как добавление к набору одной или двух хромосом может оказаться вредным.
Наиболее общее понятие о гене таково: это некая сущность с двумя основными особенностями. Во-первых, ген способен расти и делиться; во-вторых, он может вызывать изменения в протоплазме вне ядра — изменения, влияющие на химическую и физическую активность протоплазмы. Первое из этих свойств заключается в видимом делении хромосом, которые при каждом делении расщепляются продольно таким образом, что каждая половина содержит все свойства исходных хромосом. Другими словами, ген всегда делится, изменяя лишь свою массу, и затем растет, пока не достигает размеров исходного гена. Это же самое свойство должно быть у каждого саморазмножающегося нового гена. Способность деления является, следовательно, {18} свойством всей клетки, с тем лишь существенным различием, что при некоторых делениях яйца одна из дочерних клеток может содержать вещества, в некоторой степени отличные от таковых в ее сестринских клетках. Это неравенство первых делений яйцевой клетки может, как это устанавливается в последней главе, послужить нитью для пони-
 |
|
Рис. 2. Схема диспермического яйца морского ежа с 3×18=54 хромосомами и четырьмя звездами (вверху слева). Остальные рисунки показывают один из многих возможных способов распределения хромосом по четырем образующимся клеткам (по Бовери). |
мания того, почему различные области сегментирующегося яйца развиваются различными путями.
Второе свойство гена не имеет прямого опытного обоснования, но покоится на предположительно логическом выводе из результатов генетического анализа. Эти генетические данные показывают, что когда ген мутирует, то он, не теряя своего свойства саморазмножения, вызывает изменения в признаке возникающей особи. Довод этот в действительности основан на обратной связи гена с {19} признаком, так, мы обнаруживаем сначала появление нового признака или группы признаков и затем уже при помощи анализа относим это изменение признака к изменению гена. Здесь важно то, что изменение может быть аналитически прослежено до отдельной точки или локуса в одной из хромосом, т. е. до единичного гена. Предполагая, следовательно, что мы правы в заключении, что изменение в признаке зависит от каких-то свойств нового гена, возникает вопрос о том, каким образом ген влияет на протоплазму клетки, ибо именно в протоплазме проявляется признак.
Можно предполагать, что эффект гена может обусловливаться каким-либо динамическим действием гена на окружающую протоплазму. Эта возможность не может быть в настоящее время доказана или опровергнута, но так как многие или даже большинство изменений в клетках по своей природе является химическими изменениями, то кажется более правдоподобным предположить, что ген выделяет какое-то химическое вещество, — может быть вроде катализатора,— которое вызывает некоторые химические изменения в протоплазме.
Прежде чем перейти к дальнейшему рассмотрению взаимоотношения генов и признаков, необходимо еще обсудить некоторые другие вопросы. Известно, что ген при мутации вызывает изменения во всем организме. Одни из этих изменений достаточно велики для того, чтобы быть вполне видимыми, другие мельче или даже настолько мелки, что ускользают от зрительного обнаружения, проявляясь только физиологически, вызывая смерть или изменение продолжительности жизни мутанта. На заре генетики, т. е. в начале этого столетия, предполагалось, что основой для генетической работы служат «самостоятельные признаки», откуда делался вывод, что каждый ген может вызывать специфический эффект одновременно только в одном признаке. Однако очень скоро было найдено, что этот преждевременный вывод является ошибочным, так как стал известным множественный эффект каждого генного изменения. Правда, в большинстве генетических работ выбирается отдельный признак, символизирующий ген, связанный с его проявлением, но этот выбор делается только потому, что либо этот признак один из наиболее легко отличимых, либо же потому, что он менее изменчив, т. е. менее подвержен влиянию внешней среды. {20}
Следующим пунктом, требующим обсуждения, является тот факт, что каждый признак взрослого организма есть продукт действия многих генов, или можно даже сказать всех генов, если проследить до яйца всю историю развития затронутого органа.
С этими вопросами тесно связано понятие генного равновесия. Выше уже отмечалось, что эмбриологические данные показывают, что для нормального развития необходим по крайней мере один набор генов; наиболее же обычным условием является наличие двух наборов. Но известно также много случаев, в которых четыре, шесть, восемь или больше наборов тоже давали нормальные результаты. Эти факты составляют часть данных, на которых основывается понятие генного равновесия,— понятие, которое первоначально, хотя и очень смутно, подразумевалось в приведенных эмбриологических данных. Однако оно приобрело значительно более определенный смысл, когда начали учитывать отдельные гены и в свете более новых данных экспериментальной генетики прежний умозрительный характер его уступил место более определенной формулировке.
Центральной мыслью теории генного баланса является положение, что на признаки действуют все гены и что то, что получается, является результатом их общего влияния. Если изменился, т. е. мутировал, только один ген, то продукт его в какой-то степени изменяется и некоторые органы оказываются затронутыми больше других; но тем не менее все гены играют при этом роль. Другими словами, новый ген действует только как дифференциал. Эта формулировка дает удовлетворительную картину конечных продуктов генов, но совершенно недостаточна для объяснения последовательности изменений, которые претерпевает эмбрион в течение своего развития.
| << | {21} | >> |
Ни крупные яйца одних животных, ни мелкие других своим внешним видом ничего не говорят о том, какому организму они дадут начало. Каждое яйцо, хотя и имеет характерные черты, по которым может быть определен вид, к которому оно принадлежит, но эти различия являются вторичными и касаются оболочки, окружающей яйцо, присутствия пигмента, количества желтка или размеров самого яйца. Многие животные обнаруживают билатеральную симметрию, но только в редких исключениях (как например у многих насекомых) билатеральность обнаруживают неоплод отворенные яйца.
Большинство яиц имеет округлую форму; у многих из них анимальный участок отмечает определенную область, с которой связаны все начальные изменения. Вещества яйца располагаются слоями по отношению к полюсу. В анимальной области сосредотачивается больше протоплазмы, в то время как в противоположном полушарии преобладает желток (рис. 3). При взгляде со стороны полюса вещества в яйце оказываются радиально симметричными в отношении него. Какие-либо указания на то, где пройдет плоскость симметрии эмбриона, отсутствуют.
Ядро зрелого яйца исключительно велико (рис. 3) по сравнению с ядрами большинства других клеток. Оно лежит обычно эксцентрично по отношению к центру яйца, несколько ближе к полюсу, и заполнено полужидким соком, в котором находится тонкая сетка хроматинового вещества. Ядерный сок отделен от окружающей протоплазмы плотной протоплазматической стенкой.
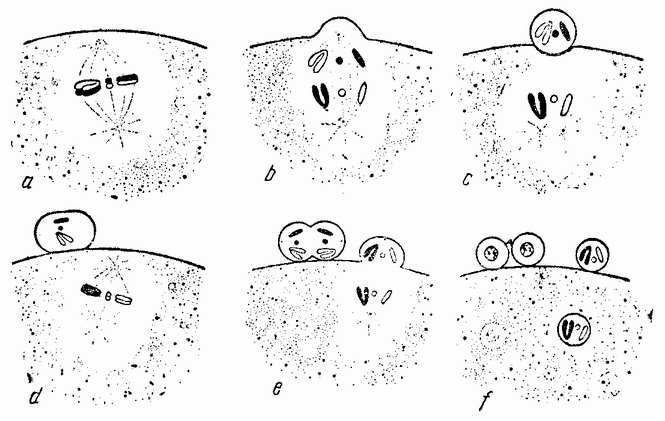
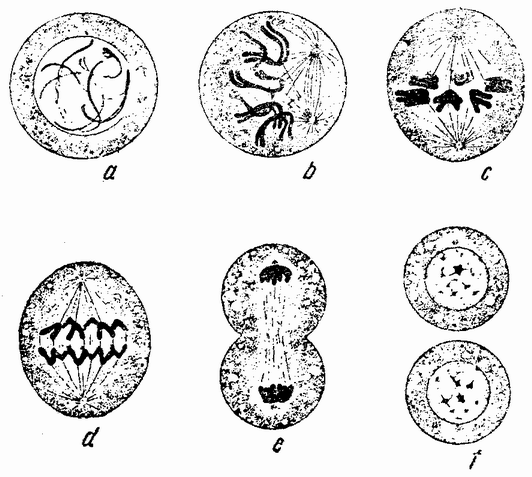
Когда яйцо готово покинуть яичник, в нем происходит соединение хромосом в пары. Стенка ядра растворяется {22} и его сок расходится по остальной протоплазме яйца. В это время появляются попарносоединенные хромосомы (рис. 4), которые располагаются в первом полярном веретене, (рис. 4, а). Число хромосом кажется здесь вдвое меньшим исходного, т. е. бывшего при более ранних делениях яйцевых клеток, так как здесь каждая хромосома состоит из двух соединившихся вместе.
Хромосомы прикрепляются к нитям веретена в его экваториальной плоскости, причем каждая хромосома имеет определенную точку прикрепления нити, которая
 |
|
Рис. 3. Зрелое яйцо лягушки. |
Веретено с хромосомами движется по направлению к полюсу яйца и принимает радиальное положение. Протоплазма на полюсе начинает выпячиваться и в образовавшийся выступ продвигается один полюс веретена (рис. 4 b). По одной хромосоме из каждой пары направляется к внешнему полюсу веретена, а их партнеры к внутреннему. Протоплазматический выступ отделяется (рис. 4 с); он содержит внешнюю группу хромосом, тогда как внутренняя остается в яйце. Выделяется первое полярное тельце.
Вокруг хромосом, оставшихся в яйце, развивается новое веретено, которое скоро принимает радиальное положение под полюсом яйца (рис. 4 d). Хромосомы, оставшиеся в яйце после выделения первого полярного тельца, в это время расщепляются, причем одна половинка каждой хромосомы {23} уходит во второе полярное тельце, а вторая половинка остается в яйце (рис. 4 е). Пока это происходит, первое полярное тельце тоже делится на две равные части. В результате возникает 4 клетки (рис. 4 f), а именно три полярных тельца и яйцо, которое произошло в результате двух клеточных делений, одно из которых отличается от всех других делений тем, что во время него хромосомы каждой пары расходятся.
Многие низшие морские животные, медузы, морские ежи, черви, моллюски и рыбы выделяют свою сперму прямо в
 |
|
Рис. 4. Деления созревания яйца; образование полярных телец. |
морскую воду, что происходит обычно одновременно с выделением яиц. Происходит потеря огромного числа сперматозоидов, но они настолько многочисленны, что все же все яйца оплодотворяются. В некоторых случаях, как например у многих саламандр, самцы откладывают сперму пачками на дно водоема, в котором в брачный период находятся оба пола. Самка ползает по этим пакетам и забирает их в клоаку. В яйцеводе пакеты растворяются, и сперма освобождается для оплодотворения яиц.
Самец лягушки обхватывает самку, влезая ей на спину, и выпускает свою сперму на яйца по мере их выхода из яйцевода. У птиц и ящериц самцы и самки совокупляются, приводя в соприкосновение отверстия клоак; сперматозоиды выбрасываются в яйцевод самки, где и происходит {24} оплодотворение. У самцов млекопитающих имеется совокупительный орган (penis), в котором проходит семяизвергательный канал. Этот орган вводится во влагалище самки, где семя испускается. Благодаря способности к движению, которой обладают сами сперматозоиды, или путем перистальтических сокращений яйцеводов сперматозоиды проходят или проводятся по яйцеводам и в их верхних отделах оплодотворяют яйца.
 |
|
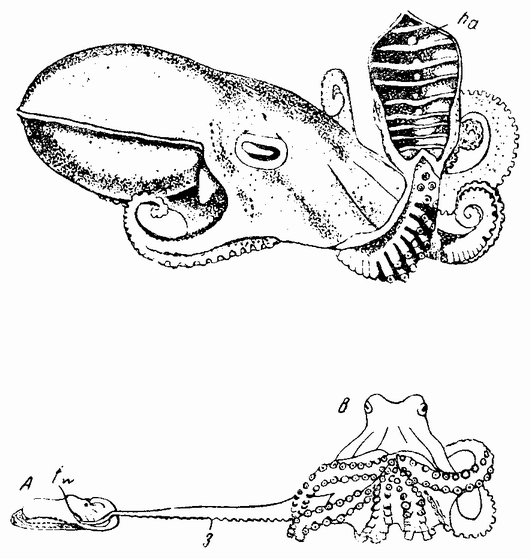
Рис. 5. Самец осьминога; виден гектокотиль (ha). Копуляция (внизу): А — мелкий самец; В — крупная самка. |
Моллюски тоже обладают совокупительными органами, при помощи которых семя переносится в яйцеводы самки. У каракатиц и осьминогов перенос пакетов сперматозоидов происходит очень сложным путем (рис. 5). Особое щупальце самца переносит пакет в мантийную полость самки и сперматозоиды освобождаются из него, как только яйца выходят из яйцевода.
У дождевого червя, являющегося гермафродитом, каждая особь образует и яйца, и сперму. Во время совокупления семя одной особи попадает в особые приемники другой особи. Непосредственно перед выходом яиц вокруг тела {25} червя позади отверстия яйцеводов выделяется поясок. Этот поясок продвигается вперед, принимает в себя яйца, и в нем появляется млечная жидкость. При прохождении отверстий семяприемника сперма извергается в поясок, который продолжает затем двигаться далее к головному концу червя и, наконец, сходит с него. У коловраток самцы вводят сперму прямо в тело самок. Отдельные сперматозоиды находят себе дорогу сквозь ткани, достигая яйца в яичнике, в котором и происходит оплодотворение. У насекомых оба пола имеют в высокой степени специализированные совокупительные органы. При совокуплении семя через penis самца проходит в яйцевод самки и попадает в ее семяприемник, где и сохраняется. При прохождении каждого яйца мимо отверстия семяприемника из него выделяется небольшое количество сперматозоидов и один или большее число их проникает в каждое яйцо. Сперматозоиды могут оставаться живыми в семяприемнике самки дни, месяцы или даже годы. Так, например, пчелиная матка, которая оплодотворяется только один раз в жизни, сохраняет достаточно живой спермы для оплодотворения яиц в течение всей оставшейся жизни, которая может продолжаться несколько лет.
В яичниках самки образуются сотни, а иногда многие тысячи яиц, которые созревают либо по одному, либо все одновременно. В яичнике молодые яйца увеличиваются в числе путем обычных процессов клеточного деления и только после многих делений они претерпевают последние стадии, в течение которых число хромосом уменьшается вдвое.
У большинства животных созревание яиц приурочено к определенному времени года, когда все зрелые яйца выводятся из организма. Животные с крупными яйцами обычно откладывают их меньше, чем животные с мелкими яйцами. Некоторые птицы каждый год откладывают только два яйца, другие дюжину или больше; лягушки откладывают несколько сот яиц; рыбы и многие низшие животные, в особенности откладывающие яйца прямо в море, выделяют большие числа их. Было подсчитано, что треска производит 6 652 000 яиц, устрица — 60 000 000, моллюск Aplysia 2 000 000. Пчелиная матка производит несколько миллионов яиц, которые она по одному откладывает в ячейки сотов в течение пяти-восьми лет своей жизни.
У млекопитающих число выделяющихся из яичника яиц относительно мало. Ежегодное число новорожденных равно {26} обычно одному у рогатого скота и лошадей и доходит до двенадцати у грызунов и свиней. Некоторые мелкие млекопитающие, как например мыши и крысы, могут давать несколько пометов в год, особенно если они содержатся в тепле и хорошо питаются. У высших обезьян и человека, у которых единственный эмбрион долгое время остается у матери в матке, общее число вырабатываемых яиц относительно очень мало, несмотря даже на то, что каждое яйцо имеет микроскопические размеры. В течение жизни женщины яичники ее могут выделить около 400 яиц, которые попадают в яйцеводы, но оплодотворяются всегда только немногие из них.
Обилие вида зависит не столько от числа вырабатываемых яиц, сколько от шансов на достижение эмбрионом взрослой стадии. Поэтому животные, защищающие свою молодь, оставляя ее в теле матери или заботясь о ней после рождения, могут оставить также много потомков, как и те, которые производят много большее число яиц.
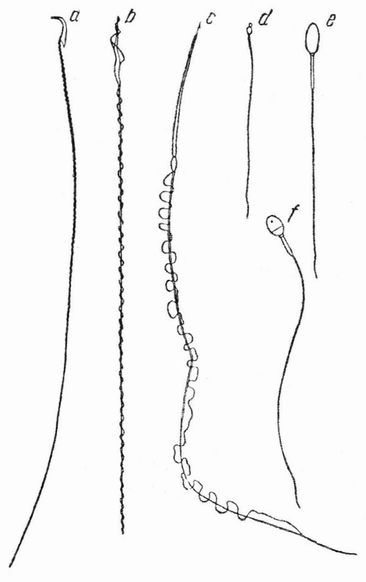
Типичный сперматозоид представляет собой мелкое нитеобразное «микроскопическое животное (анималькуль)», имеющее на одном конце округлую головку и длинный хвостик (рис. 6 а—f) Это строение может быть видимо только при очень больших увеличениях микроскопа. При попадании в воду «спермий» плавает, повидимому, самым неправильным образом, с помощью колебательных движений хвостика. Неправильность его движений, несомненно, частично зависит от столкновений с другими сперматозоидами или с мелкими частицами, плавающими в воде. Иногда можно встретить описания, что сперматозоид движется по спирали в свободной среде и циклически при соприкосновении с твердым телом.
Если в воде находятся яйца, то сперматозоиды быстро собираются в большом числе вокруг них, создавая впечатление, что они притягиваются яйцами. В действительности, однако, аккумуляция сперматозоидов является в первую очередь обусловленной прилипанием их к оболочке или к слизи, окружающей яйцо. Каждый сперматозоид, соприкоснувшийся с оболочкой яйца, прилипает к ней и через очень короткое время вокруг яйца могут оказаться сотни сперматозоидов. Прямых данных о том, что плавающие вокруг яйца сперматозоиды активно поворачивают к нему, нет, {27} наоборот, есть много указаний, что сперматозоиды могут находиться поблизости от яйца и проплывать мимо него. Большинство сперматозоидов немедленно после соприкосновения с яйцом принимает вертикальное положение,
 |
|
Рис. 6. Форма сперматозоидов: а — мыши; b — зяблика; с — тритона; d — асцидии; e — лошади; f — человека (по Балловицу, Иенсену и Броман). |
При убивании сперматозоидов соответствующими реактивами и при специальной окраске их было найдено, что они устроены весьма сложно. Впереди головки часто имеется особое тельце, называемое акросомой, функция которого хотя и неизвестна, но надо думать, что она имеет какое-то отношение к способности сперматозоида проникать сквозь оболочку яйца. Головка сперматозоида состоит главным образом из ядра той клетки, из которой он развился. Эта часть спермия состоит почти из одного хроматина, хромосомы оказываются уплотненными в плотную массу. Позади головки находится промежуточный участок, происходящий из протоплазмы исходной клетки. Он может содержать в себе одно или более особых телец, в частности клеточную центриоль или по крайней мере тельце, становящееся клеточным центром после проникновения {23} сперматозоида в яйцо. В длинном сплющенном хвостике можно иногда видеть осевую нить, окаймленную с каждой стороны плавникообразной перепонкой. Движение спермия обусловлено сокращениями хвостика, спиральный же путь его, возможно, зависит от некоторой асимметрии головки или хвостика. Проникновение сперматозоида в яйцо наблюдалось ча-
 |
|
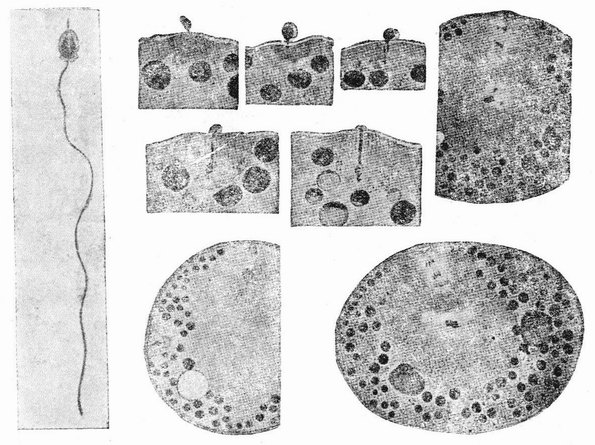
Рис. 7. Оплодотворение и образование полярных телец у Nereis. На четырех меньших рисунках изображено проникновение спермия. Выделение первого полярного тельца показано на левом нижнем рисунке, а выделение второго полярного тельца на двух больших рисунках справа. Последние три рисунка также изображают образование лучистости вокруг центриоли, внесенной сперматозоидом, которое является началом появления веретена первого дробления яйца (по Ф. Лилли). |
сто (рис. 7), но как ему в самом начале удается проникнуть сквозь часто очень вязкую окружающую оболочку — этого еще никогда не видели. Через несколько секунд наружная поверхность яйца обнаруживает явственный выступ протоплазмы в форме бугорка, находящегося как раз под головкой спермия. Как правило меньше чем в течение одной минуты головка проходит сквозь {29} оболочку, причем, когда половина ее уже прошла, она часто обнаруживает посредине перетяжку. Сейчас же после прохождения сквозь оболочку головка захватывается воспринимающим бугорком, который вместе с содержащейся в нем головкой спермия начинает в свою очередь втягиваться в яйцо.
 |
|
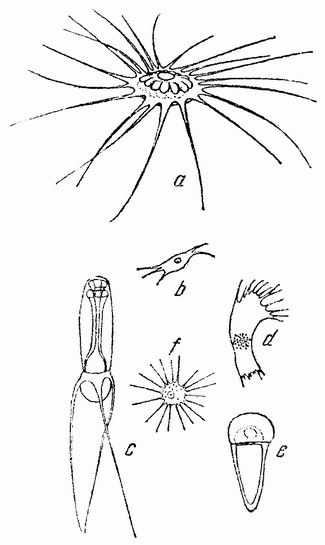
Рис. 8. Необычная форма сперматозоидов: a — речного рака; b — Polyphemus; c — омара; d — Sida; e — аскариды; f — Moina (из Коршельта и Тендера). |
Хотя большинство сперматозоидов как у низших, так и у высших групп имеет нитеобразную форму, тем не менее имеется также некоторое число других типов, лишенных хвостиков. У круглого червя — лошадиной аскариды — сперматозоид имеет амебоидную форму (рис. 8 е). Оплодотворение {30} в этом случае внутреннее — семя самца переносится к яйцевод самки. У некоторых высших ракообразных сперматозоиды имеют весьма различную форму (рис. 8 a—f), причем имеются различные приспособления, с помощью которых семя приходит в соприкосновение с поверхностью яйца.
Движение ядра сперматозоида от периферии к центру яйца происходит весьма сходным образом у всех животных.
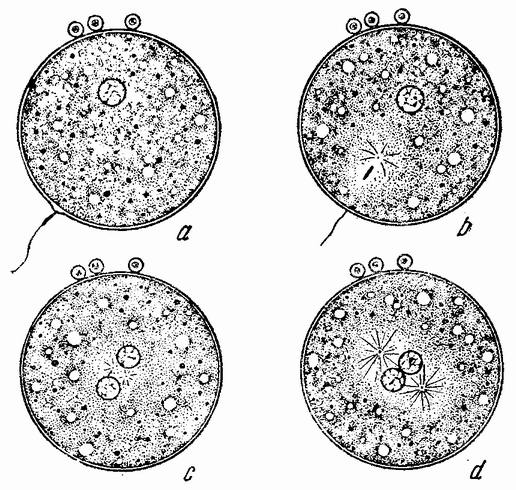 |
|
Рис. 9. Оплодотворение яйца: а — сперматозоид у поверхности яйца; b — головка проникла внутрь яйца и около нее пояиилась лучистость (звезда); с — яйцевое и семенное ядро приблизились друг к другу и звезда разделилась; d — два ядра встретились, видны два полюса митотического веретена. |
Головка сперматозоида абсорбирует из яйца жидкость и медленно увеличивает свои размеры до тех пор, пока в большинстве случаев не достигнет величины яйцевого ядра (рис. 9 с—d), которое в свою очередь тоже движется от полюса яйца к центру, где оба пронуклеуса встречаются и обычно соединяются, образуя круглое ядро.
Происхождение половых клеток — яиц и сперматозоидов — обычно может быть прослежено, начиная с ранних стадий {31} развития эмбриона. На этом факте Вейсман построил свою знаменитую теорию изоляции половых клеток, которые рано обособляются для продолжения расы, в то время как другие клетки зародыша дифференцируются для образования сомы или тела эмбриона. Половые клетки бессмертны, соматические же клетки имеют только ограниченный срок жизни. Именно сплошным рядом половых клеток связываются современные организмы с самыми древними живыми формами, тела которых давно уже истлели. Однако,
 |
|
Рис. 10. Стадии кариокинетического или митотического деления клетки. |
так как все наследуемые признаки расы содержатся в хромосомах всех клеток как половых, так и в клетках тела, то это различие больше не имеет того значения, которое ему придавалось Вейсманом. Тем не менее все же остается верным, что половые клетки как таковые обособляются также рано или даже еще раньше других клеток, дифференцирующихся в соматические органы эмбриона.
Вначале половые клетки не быстро увеличиваются в числе, но на более поздних стадиях развития эмбриона или у взрослого организма — время это весьма различно у разных форм — половые клетки делятся и число их увеличивается, что, особенно у самцов, может продолжаться в течение всей жизни. Эти первые деления половых {32} клеток, называемых оогониями и сперматогониями, происходят путем обычного процесса деления (рис. 10). Каждая клетка содержит в это время двойной набор хромосом, причем каждая хромосома расщепляется при каждом делении продольно на две равные половины и дочерние хромосомы расходятся в разные клетки. После того, как половые клетки претерпели ряд простых делений, они проходят два особых деления, в одном из которых число хромосом уменьшается на половину. Эти деления называются делениями созревания или, более специально, мейозом, в противоположность митозу или обычному процессу клеточного деления. Оба эти деления созревания яйцевой клетки происходят после того, как она накопила большее или меньшее количество желточного вещества. Эти два деления были уже описаны: они связаны с выделением двух полярных телец. Деления созревания в семенных клетках в основных своих чертах, поскольку это касается хромосом, происходят таким же образом, но из каждой материнской клетки происходит четыре функционирующих клетки вместо одной, как в случае яйца. Эти деления будут сейчас описаны.
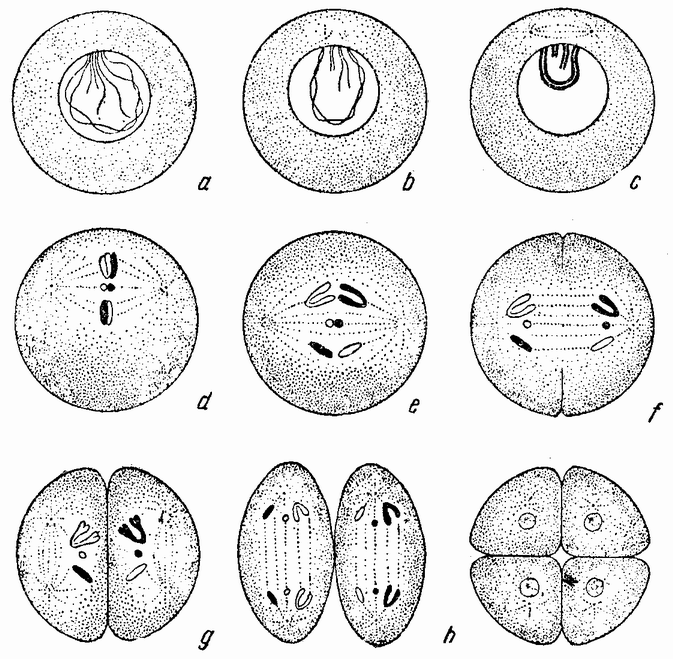
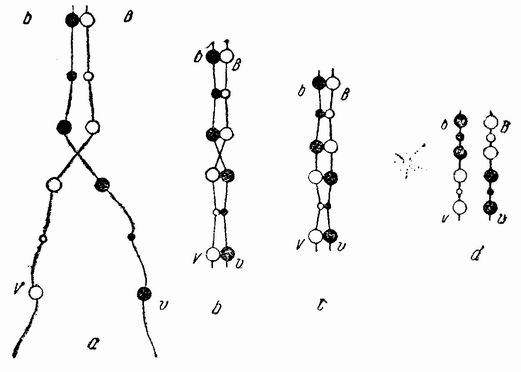
Непосредственно перед делением созревания семенных клеток хромосомы в них вновь принимают вид тонких нитей (рис. 11, a), занимающих все крупное ядро. Некоторые из хромосом имеют вид петель, другие же палочкообразны. Они располагаются так, что вдоль всей длины лежат бок о бок (рис. 11, b—с) и кажутся слившимися, но из генетических фактов известно, что в действительности они не сливаются. Является вероятным, что именно на этой стадии в яйцах или в спермиях, или в тех и в других происходит обмен между в точности эквивалентными отрезками или участками хромосом. Этот процесс называется кроссинговером (рис. 12, а—b) и в результате его возникает перекомбинация генов в гомологичных хромосомах. Далее происходит укорочение нитей и исчезновение ядерной стенки. В протоплазме появляется веретено и хромосомы прикрепляются к его нитям (рис. 11). Хромосомы выглядят еще так, как будто имеется только половина от свойственного данному виду количества, и в это время часто можно видеть, что они продольно расщеплены; иногда можно наблюдать и вторичную щель, образующую так называемые тетрады.
Затем происходит деление клетки (рис. 11, е—f—g), при котором соединившиеся пары разделяются, причем оказывается, что в некоторых случаях разделение может {33} произойти по вторичной щели или в одних хромосомах так, а в других иначе. Далее следует короткая стадия покоя и каждая клетка делится снова (рис. 11, h—i). В результате этих двух делений образуется четыре клетки, каждая из которых содержит один набор хромосом, т. е. по одной хромосоме каждого сорта. После этого следует сложный
 |
|
Рис. 11. Два деления созревания (мейоз) семенной клетки, образующих четыре сперматозоида. |
процесс диференцировки, превращающий каждую клетку в функционирующий сперматозоид.
Является очевидным, что в отношении редукции числа хромосом деление созревания яиц и спермиев в основном сходно. Было показано как наблюдениями, так и путем дедукции из генетических фактов, что разделение конъюгирующих членов каждой пары хромосом происходит случайно. Перераспределение унаследованных материнских {34} и отцовских хромосом при делении созревания имеет место и в яйцевых, и в семенных клетках.
Значение этих фактов для теории генетики лучше всего видно в тех случаях, когда особь является гибридом, то есть когда она получает от матери один гаплоидный набор хромосом, а от отца другой, причем отличный от первого. Когда яйцо близко к созреванию, хромосомы обоих наборов соединяются в пары и если (описанная выше) редукция числа хромосом происходит случайно, то конеч-
 |
|
Рис. 12. Схема, показывающая, каким образом две хромосомы, соединяясь вместе, могут перекрывать друг друга и обмениваться сегментами (кроссинговер). |
ный результат будет таков, что остающееся в яйце половинное число хромосом должно содержать в себе некоторые материнские и некоторые отцовские хромосомы, так как при случайном распределении редко все хромосомы будут оказываться только материнскими или только отцовскими. Таким образом, в каждом поколении зрелые яйца гибридной особи будут содержать все возможные комбинации хромосом, полученных от матери и отца, по одной каждого сорта. В потомстве, взятом в целом, будут все возможные комбинации прародительских хромосом. Этот процесс распределения хромосом и их соединения при оплодотворении представляет собой механизм двух менделевских законов наследственности.
| << | {35} | >> |
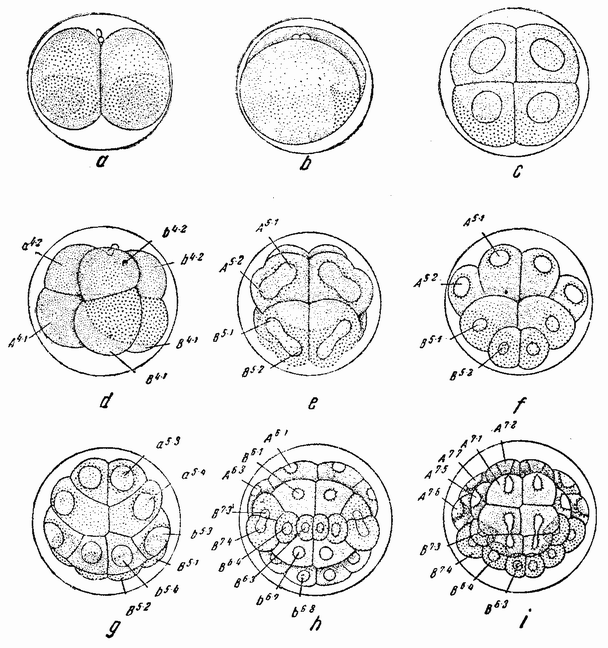
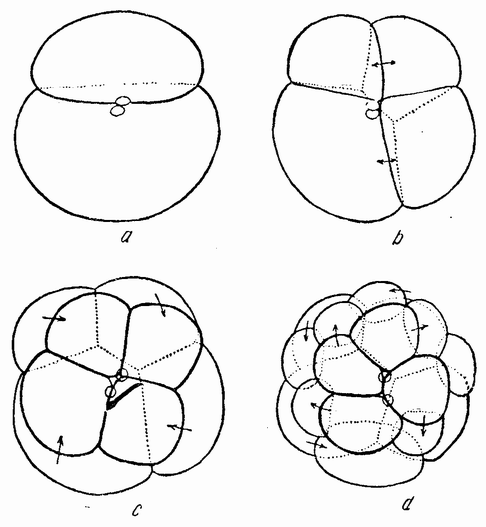
Приблизительно через час после оплодотворения яйцо начинает делиться. Деления яйца называются дроблениями. Говорят также, что яйцо сегментируется и первые образующиеся клетки часто называют бластомерами. Большинство яиц делится на две равные части, у некоторых же первое деление оказывается неравным. Оба типа являются типичными клеточными делениями в отношении поведения хромосом, веретена и т. д., но в то время как у взрослого организма последовательные деления клеток происходят только после длинного периода покоя, в течение которого дочерние клетки вырастают до размеров материнской, интервал между делениями яйца очень короткий и дочерние клетки перед следующим делением не вырастают.
Второе деление яйца следует за первым примерно через час или несколько быстрее и эта скорость остается приблизительно одинаковой до тех пор, пока не образуется большое число клеток, часто несколько сот или даже больше — тысяча, с сохранением той же массы протоплазмы, которая была вначале. Это является одним из наиболее характерных признаков дробления всех яиц. Второе деление происходит под прямым углом к первому и тоже проходит через полюс, образуя иногда четыре клетки равных размеров, но нередко две из них могут быть несколько меньше других. Третье деление происходит под прямым углом к двум первым; в типичных случаях оно лежит в экваторе или близко от него. Таким образом, первые три деления располагаются по трем измерениям пространства.
Скорость, с которой деления следуют одно за другим, является в определенных узких пределах функцией температуры. При содержании яиц на холоду скорость деления падает или даже оно полностью приостанавливается; при повышении температуры несколько выше температуры {36} нормальной среды яйца делятся быстрее, но верхний температурный предел достигается очень скоро, так как, повидимому, коагулируются некоторые важные составные части яйца. В действительности же яйца большинства видов животных очень строго «приспособлены» к средней температуре той среды, в которой они развиваются.
Среда является важным фактором в развитии яйца. Между яйцом и его внешней средой имеются постоянные взаимообмены, наиболее важным из которых является дыхание, т. е. поглощение кислорода и выделение углекислоты. Этот взаимный обмен регулируется поверхностью самого яйца, а также его оболочками. Условия на поверхности яйца играют важную роль в развитии. Состав солей, содержащихся в яйце и во внешней среде, может быть совершенно различным и это различие тоже в широких пределах регулируется поверхностной оболочкой самого яйца. Соли, содержащиеся в морской воде, играют в развитии яиц морских животных особенно важную роль. Яйца насекомых откладываются часто в сухие места; они содержат в себе все необходимые для начала развития составные части.
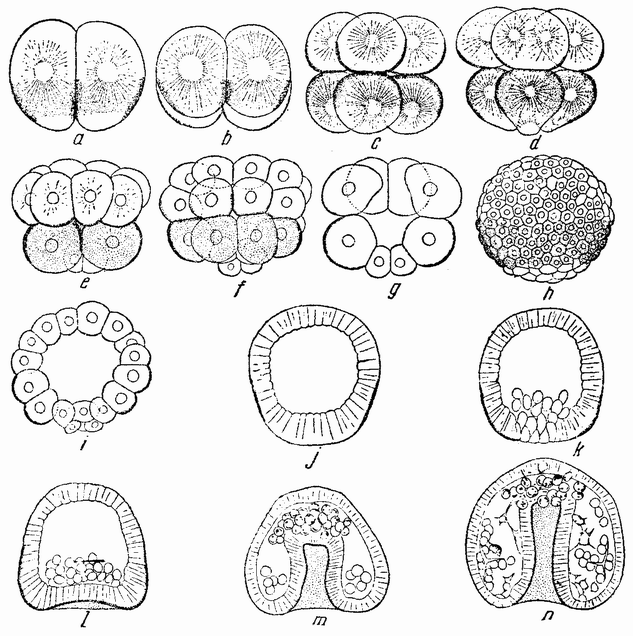
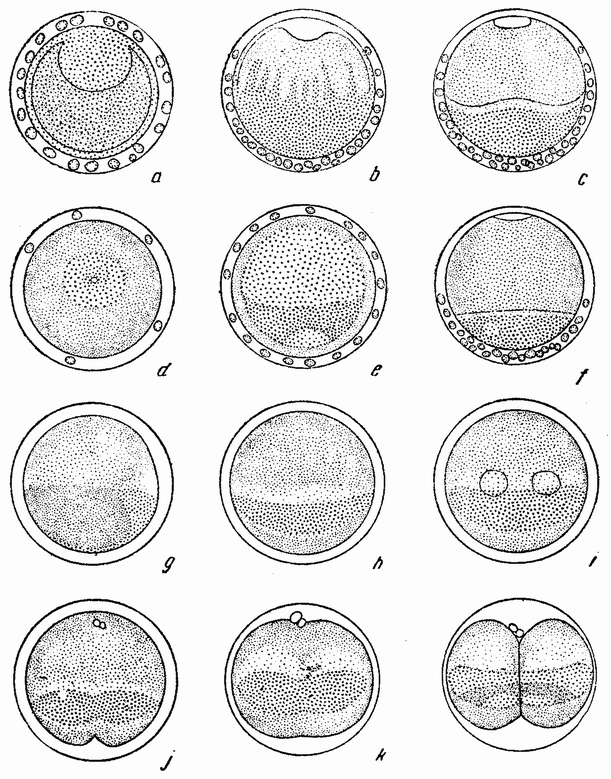
Для того чтобы глубже проникнуть в процессы, происходящие в яйце во время дробления, необходимо изучить некоторые видимые перегруппировки, которые происходят внутри яйца перед оплодотворением и сейчас же после него, еще до того как произошло первое дробление. Яйцо морского ежа, Paracentrotus, очень хорошо иллюстрирует некоторые из этих изменений. Когда яйцо еще прикреплено к стенке яичника, самый наружный слой его содержит в себе многочисленные мелкие красноватые зернышки (рис. 13, а—b), равномерно распределенные по поверхности яйца. Когда яйцо освобождается в полость яичника и стенка ядра лопается, подготовляясь к выделению полярных телец, зернышки из верхней полусферы собираются ниже экватора яйца и образуют кольцо (рис. 13, с). Это означает, что поверхностная протоплазма, в которой лежат эти зерна, передвигается из верхнего полушария яйца в нижнее. Кольцо остается в этом положении до тех пор, пока не произойдет оплодотворения и, как показано на рис. 14, а—h, продолжает существовать в течение последующих делений яйца. Наконец, при гаструляции красные клетки вворачиваются {37} внутрь яйца, становясь стенками пищеварительного тракта (14, l—n). Очевидно, что перед дроблением происходят значительные перемещения в составных частях яйца, причем имеются данные, показывающие, что такие же передвижения во время и после оплодотворения происходят и у других яиц.
Первые три борозды дробления находятся в постоянной связи с полярной осью яйца, то есть с воображаемой осью, проходящей от анимального полюса (на котором выделяются полярные тельца) через центр яйца к противоположной стороне — вегетативному полюсу. Наслоение веществ в яйце происходит под прямым углом к этой оси. Борозда пер-
 |
|
Рис. 13. Движение красного пигмента в яйце Paracentrotus во время оплодотворения (по Бовери). |
вого деления яйца морского ежа проходит через анимальный полюс (рис. 14, а), центр яйца и вегетативный полюс. Второе деление, происходящее под прямым углом к первому, тоже проходит через анимальный полюс (рис. 14, b). Третье деление проходит поблизости от экватора яйца (рис. 14, с). Восемь возникающих при этом клеток имеют приблизительно равные размеры.
Положение плоскостей дробления определяется еще до осуществления каждого деления положением веретен, которые лежат в плоскостях, перпендикулярных плоскостям будущих делений. Возникает вопрос, определяется ли плоскость деления положением веретена, где бы оно ни находилось, или же эта плоскость является фиксированной, а веретено перед делением передвигается в соответствующее положение. Данные, относящиеся к этому вопросу, получены в опытах сдавливания и центрифугирования яйц. Эти данные если и не вполне убедительны, то все же в некоторой степени дают ответ на этот вопрос. {38}
Например, путем легкого сдавливания яйца между предметным и покровным стеклами можно изменить положение плоскостей дробления; первое, второе, третье и даже более поздние дробления (рис. 15, а—е) происходят перпендикулярно к сдавливающим пластинкам. Если сплющенное яйцо окажется лежащим так, что его анимальный полюс
 |
|
Рис. 14. Дробление и гаструляция яйца Paracentrotus (по Бовери). |
будет против покровного стекла, то первые два деления пройдут через анимальный полюс и может случиться, что они окажутся лежащими на том же меридиане, на котором они были бы, если бы яйцо не было сдавлено. При этих условиях первые веретена будут лежать в своих обычных положениях. Но если яйцо окажется лежащим на боку, {39} т. е. анимальный полюс его будет под углом в 45° к сдавливающей пластинке, то положение оказывается иным; в этом случае деление пройдет все же через полюс и под прямым углом к пластинкам и так как оно проходит также и через экватор веретена, то веретено должно во многих случаях сдвигаться или образовываться на новом месте. Это более очевидно при втором делении сдавленного яйца, когда плоскость дробления вовсе не проходит через полюс, а лежит под прямым углом к нему, т. е. в первоначальной
 |
|
Рис. 15. Яйцо морского ежа, содержавшееся живым при продолжительном сжатии в струе морской воды (по Циглеру). |
экваториальной плоскости яйца, в которой проходит плоскость третьего нормального дробления. Из этого следует, что при сдавливании веретена смещаются и располагаются под прямым углом к направлению давления, т. е. параллельно стеклянным пластинкам. Следовательно, их положение, по крайней мере частично, определяет место, где пройдет борозда дробления.
Конечно, положение вещей является несколько более сложным, чем только что описанное, потому что веретено является лишь частью очень сложной митотической фигуры, занимающей почти все яйцо во время деления. Известно, что вся структура (фигура митоза) под влиянием сдавливания может сместиться и передвинуть собственно веретено {40} в новое положение, причем ответственной за этот результат является вся фигура деления, а не одно лишь веретено или прикрепленные к нему хромосомы. Как известно, фигура деления представляет собой, гель, более плотный, чем остальное вещество яйца, в котором она, так сказать, плавает. Она может быть механически сдвинута как целое и может, вероятно, также в некоторой степени быть перестроенной в своем новом положении. Наиболее убедительное доказательство этого получено при центрифугировании яиц.
 |
|
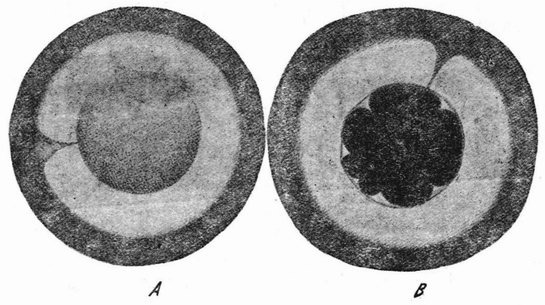
Рис. 16. Центрифугированные яйца морского ежа Arbacia. A — слоистое яйцо Arbacia, окруженное прозрачной слизью; с одной стороны видна воронка. В — восьмиклеточная стадия с микромерами напротив воронки (по Моргану и Спунеру). |
Если яйца морского ежа центрифугируются в пробирке с морской водой при очень большой скорости вращения, то они сразу опускаются на дно и более тяжелые вещества яиц перемещаются к наружному или центрифугальному краю, а более легкие к центральному или центрипетальному. Все вещества располагаются при этом четырьмя слоями, как это показано на рис. 16 А. На внутреннем конце оказывается шапка масла или жира, затем идет слой прозрачной протоплазмы, за ней широкий слой, заполненный зернышками желтка, и, наконец, слой красных пигментных зерен. Если имеется сегментационное ядро, то оно лежит в прозрачном слое непосредственно под жиром. При наличии фигуры деления она тоже располагается в этой прозрачной полосе и притом всегда параллельно слоистости, причем дробление (рис. 17, a) разрезает эту слоистость {41} под прямым углом. В студенистой массе, окружающей яйцо, имеется воронка, которая может быть сделана видимой, примешивая в воду тушь. Эта воронка соответствует анимальному полюсу яйца (рис. 16 A) и этим путем может быть показано, что в центрифугированном яйце плоскость борозды первого дробления не занимает постоянного положения по отношению к полюсу. Из этого следует, что положение плоскости дробления определяется местополо-
 |
|
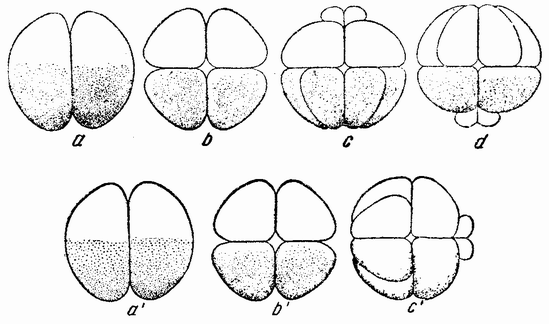
Рис. 17. Дробление центрифугированных яиц Arbacia, в которых первая плоскость деления проходит под прямым углом к слоистости, а вторая под прямым углом к первой и параллельно слоистости. Третье дробление происходит в плоскости бумаги. Четвертое дробление изображено на рис. с, d и c'; микромеры могут лежать у центрипетального полюса (с), у центрифугального полюса (d) или сбоку (с') (по Моргану). |
жением веретена. Другими словами, в нормальном яйце сама фигура деления приспособляется к положению полюса и поэтому первое деление проходит через него. Плоскость деления как таковая не предопределена анимальным полюсом или во всяком случае ее положение может быть изменено. Борозда второго деления центрифугированного яйца проходит под прямым углом к первому и параллельно слоистости (рис. 17, b), что означает, что его веретено располагается под прямым углом к веретену первого дробления. Две из образовавшихся клеток будут содержать весь жир и большую часть прозрачного вещества, в других же двух клетках будет желток и пигмент. Третье деление происходит {42} под прямым углом к первым двум, образуя четыре клетки одного рода и четыре другого.
Представляет интерес следующее дробление, при котором в точке пересечения двух плоскостей деления возникают четыре микромера (рис. 17, с—d). Они могут лежать в месте пересечения плоскостей первого и второго или первого и третьего деления в пигментном или в желточном полях, или между ними. Было найдено, что где бы они ни располагались, микромеры образуются напротив воронки или возможно более близко от нее. Из этого, повидимому, следует, что ко времени появления микромеров в яйце имеется область, дающая им начало, и что образование их при четвертом дроблении в высокой степени независимо от последовательности появления предыдущих плоскостей.
На более поздней стадии микромеры перемещаются внутрь бластулы (рис. 14, k—n) и превращаются в мезенхиму. Может казаться поэтому, что в яйце имеется определенная область, которой предначертано стать микромерами; но центрифугирование яиц не дает ответа на вопрос, локализуется ли эта область на вегетативном полюсе до или во время выделения полярных телец или, может быть, она образуется где-либо в другом месте и передвигается в определенную часть яйца после того, как оно уже оплодотворено или во время первых дроблений. Имеются, однако, другие опыты, относящиеся к этим вопросам.
Если яйца морских ежей сильно встряхнуть в небольшой пробирке, то они могут быть разломаны на части, которые затем могут быть оплодотворены. Некоторые из этих частей будут содержать пронуклеус яйца, в других же он будет отсутствовать, но как те, так и другие при проникновении в них сперматозоида будут развиваться. Образование микромеров в фрагментах очень неправильно и так как фрагменты являются различными и неизвестными частями яйца, то из этих результатов не может быть сделано достоверных выводов. Более точный метод заключается в разрезании яиц на две части и в прослеживании развития каждой из них. К сожалению, в большинстве яиц морского ежа пронуклеус перед оплодотворением не занимает определенного фиксированного положения по отношению к анимальному полюсу; полярные тельца теряются и воронка не может быть видна без обработки тушью. Но у вида Lytechinus полярные тельца сохраняются, что делает возможным ориентировку разреза. Если эти яйца разрезаны в {43} вертикальной плоскости, т. е. вдоль оси яйца, то после оплодотворения развиваются оба куска, один из которых оказывается гаплоидным, а другой диплоидным. Однако оба эмбриона (плутеуса) не могут быть отличены от нормальных, кроме как по своим размерам. Когда оба фрагмента делятся, внешний вид одного из них кажется нормальным, другой же фрагмент не имеет микромеров. Это, повидимому, означает, что либо разрез прошел с одной стороны оси и вегетативный полюс находится только в одном фрагменте, либо же, что вещество микромеров было в это время в центре или вблизи центра яйца и лежит с одной стороны разреза. Последующие опыты делают трудным окончательное решение, так как исследователи пришли к заключению, что в этих яйцах отсутствует локализованное вещество, образующее микромеры, — по крайней мере такое вещество не «диференцируется» перед оплодотворением. Что такое вещество может тем не менее присутствовать в яйце даже если оно и не диффузно распространено на поверхности или сконцентрировано вблизи центра яйца, не было убедительно показано. Однако существуют другие опыты, проведенные на яйцах морского ежа Arbacia, показывающие, повидимому, что такое вещество присутствует внутри яйца еще до того, как произошло оплодотворение. В связи с этим надо напомнить, что в яйцах Arbacia вещество, из которого образуются микромеры, всплывает изнутри яйца к вегетативному полюсу, когда веретена микромеров касаются этого участка. В действительности микромеры состоят почти полностью из вещества внешней полярной звезды и только тонкий поверхностный слой состоит из исходного анимального вещества.
Со времени первых наблюдений Эдварда Ван Бенедена над яйцами асцидий они являлись излюбленным объектом для экспериментальной работы. Некоторые из этих яиц прозрачны, как стекло, другие же содержат пигментные зерна, движения которых служат прекрасными отличительными признаками, очень же ранняя диференцировка клеток дает возможность проследить их до конца. У этого объекта имеется только один недостаток, заключающийся в вязкости оболочки вокруг яйца, а также в наличии между поверхностью яйца и оболочкой внутреннего кольца особых клеток (рис. 18, а) и второго слоя {44} их над наружной поверхностью оболочки. Недавно был найден метод растворения оболочки без повреждения яйца.
 |
|
Рис. 18. Движение веществ в яйце асцидий Styela во время оплодотворения (по Конклину). |
Зрелые яйца покидают яичник и собираются в яйцеводе. Полярное веретено в это время уже присутствует, но {45} полярные тельца не выделяются до тех пор, пока сперматозоид не проник в яйцо. Сперматозоид проникает в яйцо на вегетативном полюсе или очень близко от него. У Styela яйца содержат в поверхностном слое зерна желтого пигмента (рис. 18, а). Когда сперматозоид проникает в яйцо, внешний слой протоплазмы быстро скользит по поверхности, увлекая с собой окрашенные зерна (рис. 18, b). На одной стороне образуется желтый полумесяц, занимающий более полуокружности яйца (рис. 18, h—l). Этот полумесяц является первым указанием на билатеральный характер яйца.
Плоскость первого деления проходит через середину полумесяца (рис. 18 l), причем половина его попадает в одну клетку, а половина в другую. Попадает ли поверхностный протоплазматический слой и желтый пигмент полумесяца на предетерминированную сторону яйца или же их положение определяется точкой проникновения сперматозоида — до сих пор еще спорно. Однако одно, повидимому, достаточно хорошо установлено: головка сперматозоида двигается сначала в ту сторону, где находится середина полумесяца. Если сперматозоид проникает в яйцо не точно в вегетативном полюсе, а несколько в сторону от этой точки, а это, повидимому, является правилом, то эта эксцентрическая точка может служить детерминирующим фактором, приводящим к более экстенсивному накоплению поверхностной протоплазмы (и пигмента) на этой же самой или на противоположной стороне яйца. Если это рассуждение правильно, то билатеральное распределение содержимого яйца не предетерминировано, а является вторичным и, как и в некоторых других яйцах, обусловлено точкой проникновения сперматозоида.
Первое деление яйца проходит через анимальный полюс и через середину полумесяца (рис. 19, а—b), разделяя яйцо на две в точности равные части. Второе деление тоже проходит через полюс и под прямым углом к плоскости первого деления. Две из образовавшихся клеток несколько крупнее других двух и содержат большую часть полумесяца (рис. 19, с). Две меньших клетки содержат только рожки полумесяца и могут быть названы передними клетками. Протоплазма передних и задних клеток также несколько отличается по своему виду и, несомненно, их содержимое отличается и в других отношениях, а не только видимым количеством содержащегося в них пигмента или желтка. {46}
Борозда третьего деления проходит под прямым углом к первым двум и отрезает от каждой клетки (рис. 19, d) меньшую клетку на анимальном полюсе и более крупную на противоположном конце яйца. Две из последних содер-
 |
|
Рис. 19. Стадии дробления яйца Styela (Cynthia) partita (по Конклину). |
жат большую часть желтой полосы полумесяца. Начиная с этого времени, деления продолжают происходить обычным образом, образуя клетки, которые могут быть идентифицированы по своему содержимому и местоположению. Судьба этих клеток будет описана в главе о гаструляции. {47}
Некоторые опыты сдавливания, проведенные на яйцах асцидий, показывают, что и эти яйца, подобно другим, делятся под прямым углом к сдавливающим пластинкам; но в отличие от сдавленных яиц морского ежа ни одно из яиц асцидий при освобождении от сдавливания не образует целиком нормального зародыша, хотя бы даже они и продолжали делиться. Некоторые из таких яиц дают начало зародышам с перемещенными органами. Это объясняется проще всего тем, что в этих очень рано диференцирующихся яйцах любое нарушение правильного распределения веществ приводит к катастрофическим последствиям.
Изучалось также влияние центрифугирования на эти яйца. Благодаря большей вязкости протоплазмы у таких яиц труднее вызвать слоистость, чем у многих других. Если яйца Styela центрифугируются перед оплодотворением, то желтый пигмент может быть сдвинут к одному полюсу; поверхностная протоплазма предположительно претерпевает свои нормальные движения и эти яйца могут образовать нормальных зародышей, причем пигмент может оказаться лежащим в любой части эмбриона. Это доказывает, что желтый пигмент как таковой не является органообразующим. Тем не менее его передвижение в нормальном яйце говорит об изменениях в перераспределении протоплазматического содержимого яйца, которые связаны с последующими влияниями на развитие. Имеются данные, показывающие, что у других асцидий, не имеющих пигмента, например, у Ciona, так же как и у Styela, происходят передвижения поверхностной протоплазмы, приводящие к образованию полумесяца.
Благодаря вязкой оболочке трудно получить фрагменты яйца асцидин, происходящие из определенных участков, и до последнего времени было невозможно отрезать фрагмент яйца из определенной области его. Тем не менее было показано, что фрагменты делятся и пытаются образовывать нормальные зародыши, но до тех пор, пока не будет точно определена связь фрагментов с определенной областью яйца, рискованно обобщать эти неадекватные результаты.
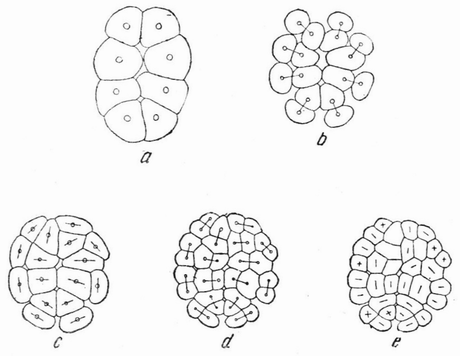
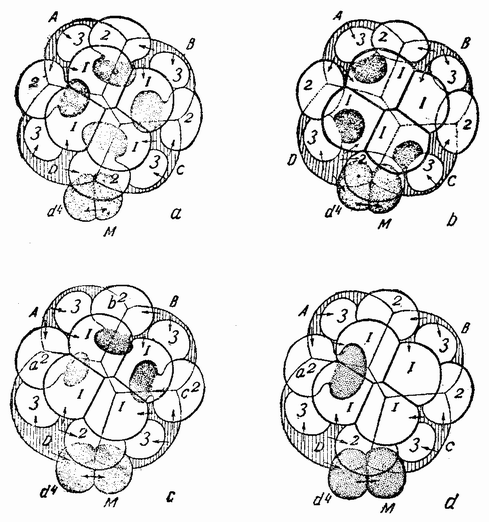
Хорошо известны и другие типы дробления, отличные от таковых у морского ежа и асцидий. Спиральный тип дробления аннелид и моллюсков отличается от предыдущего {48} типа главным образом тем, что при третьем дроблении вокруг полюса обособляются четыре клетки, микромеры, которые образуют вправо или влево закрученную спираль (рис. 20). При шестом дроблении на экваторе закладывается особая асимметричная клетка, называемая d4, дающая начало мезодерме или среднему слою зародыша
 |
|
Рис. 20. Поздние стадии дробления яйца планарии (a), аннелиды (b) и двух моллюсков (c и d). Показано положение клетки d 4 (no Вильсону). |
(рис. 20). Ее последующее деление на две маркирует первую резко различимую билатеральную плоскость зародыша.
Самки морской аннелиды Nereis всплывают иногда в летние ночи на поверхность моря и очень быстро плавают. Одновременно подымаются на поверхность и самцы. Если один из них приближается или сталкивается с самкой, то она немедленно выбрасывает все свои яйца, а самец одновременно изливает свою сперму. Яйца оплодотворяются {49} в морской воде. Каждое яйцо прозрачно и при выделении содержит крупное яйцевое ядро, которое быстро исчезает при оплодотворении, освобождая свои хромосомы. Яйцо окружено толстой оболочкой с радиальными порами. Под оболочкой имеется слой, содержащий жидкое вакуолизированное вещество, которое сразу проходит через оболочку, как только сперматозоид касается поверхности яйца.
 |
|
Рис. 21. Дробление яйца Nereis. Образование первой (с) и второй (d) четверки микромеров. На рис. d видно деление первых микромеров (по Вильсону). |
Жидкость коагулируется в морской воде и становится желеобразной, покрывая собой яйцо. Она продолжает в течение нескольких часов набухать до тех пор, пока диаметр яйца вместе с окружающей его слизью не станет по крайней мере в три раза больше диаметра первоначального.
Проникновение сперматозоида через оболочку яйца продолжается более получаса. Его головка втягивается {50} выступающим бугорком оплодотворения; хвостик сперматозоида остается снаружи и служит маркером, с помощью которого может быть определено положение плоскости первого дробления по отношению к точке проникновения спермия. В большом проценте случаев они совпадают полностью и приблизительно совпадают в большинстве остальных случаев. Два полярных тельца выделяются внутри оболочки. Первое дробление (рис. 21, а) делит яйцо на несколько неравные части. Борозда второго дробления, проходящего под прямым уголом к первому, делит меньшую клетку на две равные, а большую на две неравные части (рис. 21, b). В результате вокруг оси яйца лежат три более мелких клетки приблизительно равной величины и одна более крупная клетка.
При третьем дроблении ясно обнаруживается спиральное деление яйца. Наискосок вокруг полюса обособляются четыре мелких клетки (рис. 21, с), каждая слева от своей сестринской клетки. Каждая клетка содержит очень прозрачную протоплазму, не имеющую желточных и жировых шаров, имеющихся в других клетках. Четыре микромера лежат в промежутках между четырьмя крупными клетками, причем из косого расположения веретен еще до того, как микромеры начинают отщепляться от крупных клеток, видно, что это расположение является не простым механическим заполнением полезного пространства после деления. При следующем, четвертом, дроблении (рис. 21, d) этот же самый процесс повторяется, но в это время веретена в крупных клетках поворачиваются в другом направлении, то есть вправо, давая начало второй четверке микромеров. Первые микромеры делятся, образуя спираль, закрученную в противоположную сторону. При пятом дроблении слева образуются еще четыре микромера — третья группа их. В это время справа добавляется четвертая группа микромеров из трех крупных клеток и четвертого микромера (d1), который является много более крупной клеткой (рис. 20, b), лежащей вдоль оси исходного второго деления. Эта клетка является предшественником большинства мезодермальных клеток зародыша. При ее делении на две равные клетки определяется средняя линия зародыша, проходящая между ними. Эта средняя линия не соответствует в точности какому-либо из первых двух делений, но очень близка к плоскости борозды второго деления. Микромеры продолжают делиться и, наконец, покрывают поверхность эмбриона. {51} Крупные клетки, содержащие желток, дают начало первичному кишечнику.
Деление яиц большинства моллюсков (рис. 20, c—d) за исключением каракатиц и осьминога, которые имеют крупные яйца, в принципе и часто даже в большинстве деталей, подобно делению яйца Nereis, несмотря на тот факт, что взрослые животные, принадлежащие к этим двум большим группам, совершенно различны. Все же сходство
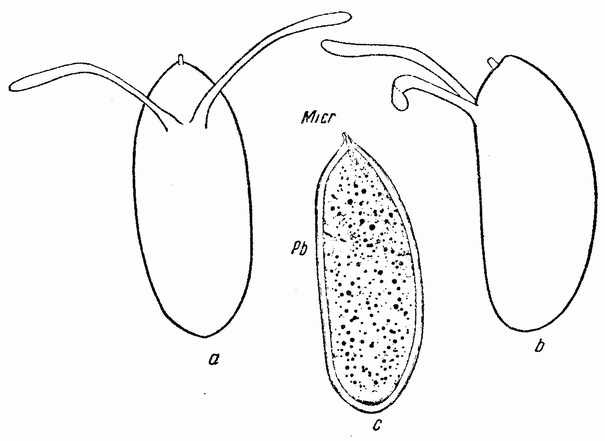 |
|
Рис. 22. а — билатеральное яйцо дрозофилы с дорзальной стороны; b — с правой стороны; с — разрез яйца через среднюю плоскость и микропиле (Micr). Полярные тельца (Pb) выделились в верхней трети дорзальной стороны. |
самых первых стадий и тот факт, что происхождение их органов так сходно, вряд ли может оставлять сомнения в том, что эти две группы родственны и что в далеком прошлом их общие ископаемые предки, о которых мы не имеем никаких сведений, проходили через стадии развития, подобные обнаруживаемым у представителей обоих этих типов, живущих в наши дни.
Яйца насекомых развиваются совершенно другим путем. Если предками насекомых были, как это принято думать, аннелиды, то они приобрели резко отличный тип {52} дробления. Яйца большинства насекомых имеют не сферическую форму, как это имеет место у большинства морских и пресноводных животных, а продолговатую, причем форма эта фиксируется на последних стадиях развития яиц и зависит от формы яйцевых трубок, в которых они образу-
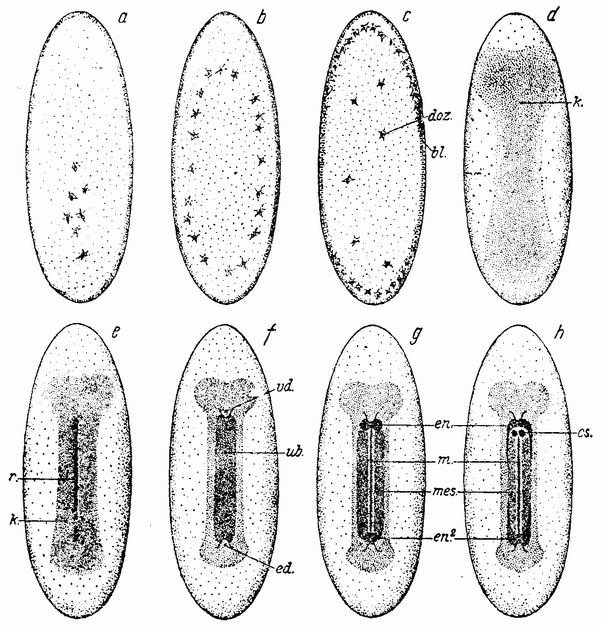 |
|
Рис. 23. Дробление и образование зародыша у насекомого (по Мангольду). |
ются (рис. 22, а, b). Форма яиц обеспечивается очень твердой окружающей их оболочкой, которая пронизана одной или многими порами, находящимися не на конце яйца, а на одной из его сторон. Многие яйца билатеральны, причем одна сторона у них более плоская, чем другая (рис. 22, b—с). Стороны яйца соответствуют вентральной и дорзальной {53} сторонам эмбриона. Яйцо имеет билатеральную форму, когда оно готово к откладке. При продвижении яйца через заднюю часть яйцевода микропиле соприкасается с отверстием семяприемника, мимо которого оно проходит. В яйцо попадает несколько сперматозоидов, которые проникают в него вместе с хвостиками. Яйцо выделяет полярные тельца и яйцевое ядро глубже погружается в протоплазму. Ядра сперматозоидов движутся по направлению к ядру яйца и одно из них приходит в соприкосновение с последним. Тогда остальные ядра сперматозоидов пре-
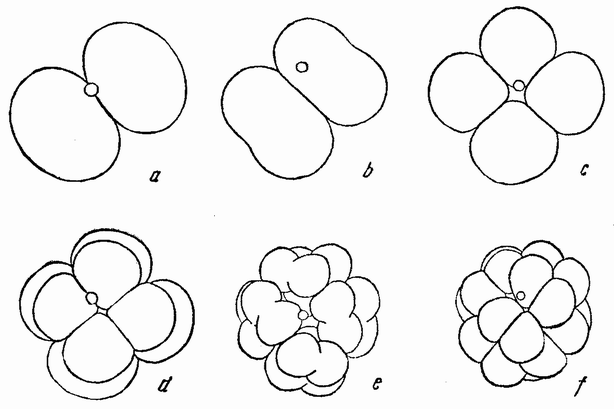 |
|
Рис. 24. Первые четыре дробления яйца ланцетника (по Церфонтену). |
кращают двигаться к яйцевому ядру и впоследствии дегенерируют.
Внутренность яйца выполнена желточными гранулами, заключающимися в протоплазме; поверхность же яйца покрыта слоем протоплазмы, свободной от желтка (рис. 23, а). Веретено образуется поблизости от слившихся ядер и вокруг него собирается небольшое количество протоплазмы. Стенки ядра исчезают и хромосомы входят в веретено, где они делятся и расходятся к полюсам. Получается два ядра, однако протоплазма не делится. Эти ядра делятся опять, причем процесс этот продолжается до тех пор, пока число образовавшихся ядер не станет очень велико (рис. {54} 23, а—b). После каждого деления ядра расходятся в стороны и скоро образуют кольцо или даже сферу (рис. 23, b). По мере увеличения их числа, ядра приближаются к поверхности яйца и, наконец, оказываются лежащими в поверхностном слое протоплазмы (рис. 23, с). К этому времени имеется
 |
|
Рис. 25. Каждая пара рисунков изображает противоположные стороны яйца лягушки во время первых дроблений. Более темная сторона на левых рисунках — так называемая вентральная сторона яйца, а более светлая сторона на правых рисунках — так называемая дорзальная сторона или сторона серого полумесяца. На рис. е и f изображены клетки дорзальной стороны, которые мельче таковых на противоположной стороне яйца на том же самом уровне (по Браше). |
уже несколько сот ядер. Далее, поверхностная протоплазма начинает дробиться, причем каждая борозда проходит между половинками делящегося ядра. Эти борозды располагаются радиально, но не доходят до внутренней стороны {55} наружного слоя протоплазмы, оставляя каждую клетку открытой на своем внутреннем конце; содержимое клетки и течение некоторого времени оказывается слитым с центральной желточной массой. На заднем конце яйца одно или несколько ядер переходят в особую протоплазму; эти клетки являются предшественниками половых клеток.
Яйца низших ракообразных заключены в твердую скорлупу. Они делятся на два, четыре, восемь и т. д. клеток, причем у речного рака и омара обнаруживают почти такой же тип дробления, как и у насекомых. У них имеется центральная желточная масса и поверхностный слой клеток, перегородки между которыми продолжаются до центра яйца.
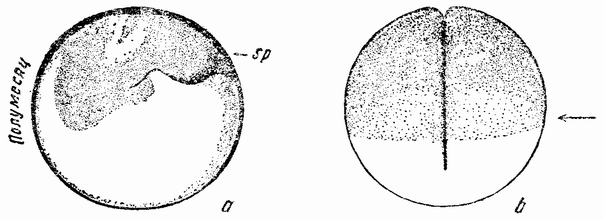 |
|
Рис. 26. а — разрез через яйцо лягушки, через полумесяц в плоскости первого дробления. На стороне яйца, противоположной полумесяцу, виден след проникшего сперматозоида (sp); b — вид с поверхности на делящееся яйцо лягушки. Дробление разрезает полумесяц (по Шульце). |
У позвоночных имеется два основных типа дробления. Ланцетник, являющийся низшим членом группы, имеет мелкие яйца, которые делятся сначала на две равные части, затем на четыре, восемь и т. д. (рис. 24, а—d), почти так же, как у асцидий. Яйцо лягушки, которое много крупнее, тоже имеет полное дробление (рис. 25, а—f). В первых стадиях развития этих яиц имеется одна особенность, которая напоминает развитие яйца асцидий. Как только в верхнее полушарие яйца проник сперматозоид, на той стороне яйца, которая лежит против точки проникновения сперматозоида, происходит движение поверхностного вещества. Серый полумесяц появляется вдоль края между черными и белым полушариями (рис. 26, a, b). Середина полумесяца, как и у асцидий, становится средней плоскостью зародыша. {56} Через его среднюю линию, как правило, проходит первое деление; но независимо от того, проходит ли это деление через середину полумесяца или нет, оно изредка может быть на 90° под углом к нему — средняя плоскость зародыша соответствует середине полумесяца.
Большинство яиц костистых рыб мельче, чем яйца лягушки, но тем не менее не имеют полного дробления. Их яйца состоят из центральной желточной массы, покрытой очень
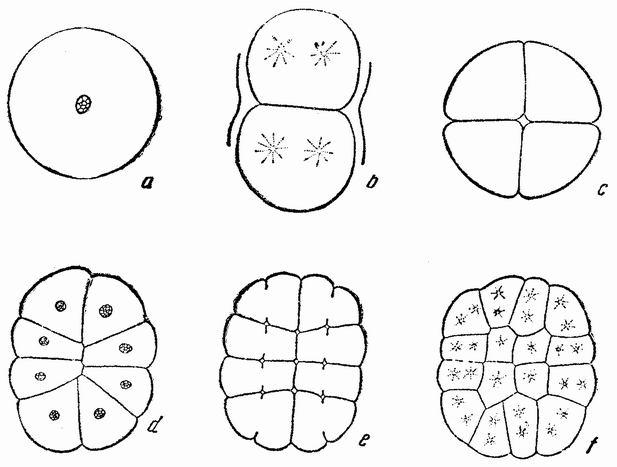 |
|
Рис. 27. Первые четыре дробления яйца костистой рыбы (по Г. В. Вильсону). |
тонким слоем или шапкой протоплазмы, которая несколько толще над анимальным полюсом. У полюса через окружающую яйцо оболочку проходит небольшая трубка, по которой проникает сперматозоид. Первое дробление (рис. 27, b) состоит из прямой борозды в полярной шапке протоплазмы; борозда второго дробления проходит под прямым углом к первому; борозда третьего под прямым углом ко второму (рис. 27, с, d). Положение следующих делений несколько неправильно (рис. 27, е, f). Первая борозда сначала не сильно углубляется в яйцо; после {57} образования нескольких поверхностных бороздок самые центральные клетки делятся под прямым углом к последним, образуя внешнюю клетку и оставляя внутреннее ядро в поверхностном желтке.
Яйца рептилий и птиц имеют относительно огромную величину. Им свойственен желток, вокруг которого, по мере прохождения яйца по яйцеводу, откладывается слизь («белок»). Еще ниже в яйцеводе вокруг яйца образуется известковая скорлупа. Оплодотворение происходит, как только яйцо покидает яичник и входит в яйцевод. Дробле-
 |
|
Рис. 28. Сегментация яйца млекопитающего, f и g — начало образования бластоцисты и бластодермы (по Прентису). |
ние заключается сначала в образовании поверхностных бороздок, как и у яиц рыб. Образуется зародышевый диск, состоящий из многих клеток и лежащий на анимальном полюсе яйца.
Млекопитающие произошли от форм, имевших крупные яйца, подобные таковым рептилий. Низшее из живущих в настоящее время млекопитающих, Platypus, еще сохранило этот тип яйца, размеры которого три миллиметра в диаметре, но яйца всех других млекопитающих почти не имеют желтка, вследствие чего они очень мелки и едва видимы невооруженным глазом. Яйцо млекопитающего полностью делится на две, четыре, восемь клеток почти {58} равной величины (рис. 28, а—с). Внутри клеточной массы появляется жидкость (рис. 28, f), и клетки покрывают стенки образующегося пузырька (рис. 28, g), который постепенно увеличивается до значительной величины (рис. 29, a, b). В одной точке несколько раньше образовавшихся клеток соскальзывают под внешний слой, который затем покрывает
 |
|
Рис. 29. Развитие млекопитающего, а — бластоциста с бластодермой сверху; b — более поздняя стадия с эндодермой, образующей «желточный» мешок; с — зародыш более поздней стадии с «желточным» мешком и плацентой; d — вид молодого эмбриона с поверхности (из Кибель и Маль). |
их (рис. 29, а). Внутренняя группа клеток образует тогда небольшой диск, который затем дает начало эмбриону (рис. 29, d). Стенки пузырька соединяются со стенками {59} матки материнского организма (рис. 29, с), в которой зародыш претерпевает свое дальнейшее развитие.
В развитии млекопитающих имеются две стадии, которые казались бы непонятными, если не знать, что млекопитающие произошли из форм, имевших крупные яйца.
Молодые клетки, которые впоследствии станут яйцами, сначала лежат в поверхностном слое яичника (рис. 30, а).
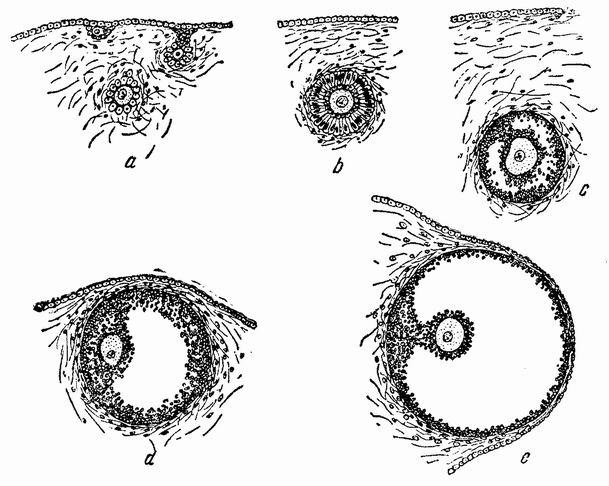 |
|
Рис. 30. Четыре стадии яйца и граафова пузырька у млекопитающего (из Пэтена). |
Некоторые из них погружаются внутрь яичника, увлекая с собой группу других клеток, которые окружают каждое яйцо. По мере роста яйцевых клеток между клетками, непосредственно окружающими яйцо, и внешним слоем клеток появляется жидкое пространство (рис. 30 с, d, e); это пространство становится относительно огромным и на поверхности яичника выпячивается в виде шара, называемого граафовым фолликулом. Если бы не присутствие жидкости в фолликуле, то он напоминал бы крупный фолликул яичников птиц и ящериц, у которых яйцо, содержащее {60} желток, целиком заполняет внутренность фолликула. Если мы истолкуем величину фолликула как напоминание о состоянии предков, когда яйца млекопитающих содержали желток, то мы будем иметь своего рода объяснение современного состояния. Это объяснение постулирует, что яйцо млекопитающего сохранило в настоящее время некоторые из стадий развития своих предков.
Когда яйцо созревает, фолликул лопается. Воронкообразное отверстие яйцевода, открывающееся в полость тела, захватывает из яичника яйцо, которое проходит по переднему отделу трубы, и затем закрепляется в стенке
 |
|
Рис. 31. а — эмбрион и желточный мешок птицы; b — эмбрион и желточный мешок млекопитающего (а — по Лилли). |
матки, где и происходит его дальнейшее развитие.
Другая характерная стадия развития заключается в образовании желточного мешка, в котором отсутствует желток. Когда на бластодерме почти образовался зародыш (рис. 29, b), края энтодермы тянутся вокруг внутренней стенки эктодермального пузырька, образуя внутренний шар или мешок; этот мешок съеживается и отстает от наружной стенки и по мере вклинивания между ними мезодермы образует «желточный мешок» (рис. 31, b), основание которого связано со стенкой кишечника зародыша, и становится пупком. Впоследствии этот мешок втягивается в стенку пищеварительного тракта. Эмбрион, так сказать, «обманул» желточный мешок, хотя последний и не содержит в себе желтка. Другими словами, млекопитающее развивается так, как будто бы оно произошло из крупного, содержащего желток, яйца, подобного яйцу птицы (рис. 31, а) {61} или рептилии, которое следует еще способу развития своих предков. Желточный мешок является рудиментарным органом или эмбриональным атавизмом. Другими словами, генный комплекс, играющий роль в этой стадии развития, не был изменен. Конечно, проще предположить, что эти стадии зависят от особых генов, действие которых не затрагивалось изменениями в других генах, вызывавших дальнейшие эволюционные стадии.
| << | {62} | >> |
Процесс, называемый гаструляцией, заключается в инвагинации наружной стенки полого шара или бластулы для образования пищеварительного тракта. Наиболее простая форма гаструляции наблюдается при развитии морского ежа (рис. 14, l—n) или ланцетника (рис. 33, а—с), а наиболее модифицированная — при развитии птиц и млекопитающих.
В конце периода наиболее быстрого деления клеток яйца морского ежа образуется полый шар (бластула) (рис. 14, i—j), стенка которого состоит из единственного слоя коротких цилиндрических клеток. Эти клетки мельче в анимальном полушарии и крупнее в вегетативном. У Раracentrotus вблизи экватора, но главным образом в вегетативном полушарии, лежит пигментное кольцо. Группа микромеров мигрирует внутрь бластулы для образования мезенхимных клеток (рис. 14, k—n), из части которых вскоре развивается личиночный скелет. Затем слой клеток нижнего полушария вворачивается, т. е. инвагинирует, внутрь, образуя подобие трубки, называемой первичным кишечником, который тянется почти до противоположной стенки гаструлы.
Из самого внутреннего конца трубки образуется выступ, который перетягивается на два мешка, образующие полость тела и гастроваскулярную систему личинки. Далее внутренний конец первичного кишечника загибается на одну сторону и приходит в соприкосновение с внутренней стенкой бластулы. В точке соприкосновения стенка бластулы инвагинирует внутрь в виде небольшого мешка или короткой трубки, становящейся ртом и пищеводом личинки.
Стадия бластулы, по всей вероятности, уже с самого начала радиально симметрична вокруг полярной оси. Видимых признаков будущей билатеральной структуры у нее {63} нет, но имеются некоторые экспериментальные данные, показывающие, что в большом проценте случаев билатеральность имеется, причем плоскость симметрии бластулы соответствует плоскости борозды первого дробления. Имеется ряд указаний на то, что эта связь существует, но наиболее доказательные данные были получены с помощью прижизненной окраски. Яйца на стадии двух клеток приводились в соприкосновение с маленькими пластинками
 |
|
Рис. 32. Способ окраски яйца морского ежа на двухклеточной стадии, а и b — с помощью кусочка агара, окрашенного «нильблау»; с—f — более поздние стадии гаструлы и молодой плутеус, развившиеся из таких яиц (по Убишу). |
агар-агара, которые были предварительно намочены в ниль-блау (рис. 32, а—b). Часть яйца, соприкасавшаяся с пластинкой, становилась синей. Когда один из бластомеров частично окрашивался, яйцо удалялось. Большинство развившихся эмбрионов было окрашено с одной стороны, показывая этим, что плоскость первого дробления совпадает с средней плоскостью зародыша. Однако в некоторых случаях окрашенной оказывается передняя или задняя {64} сторона эмбриона, что доказывает либо непостоянство этой связи, либо возможность изменения ее (рис. 32, с—f). У ланцетника процесс гаструляции (рис. 33, а—с) весьма сходен с гаструляцией морского ежа, за исключением лишь того, что инвагинированная часть (первичный кишечник) более широко открыта, образуя гаструлу, имею-
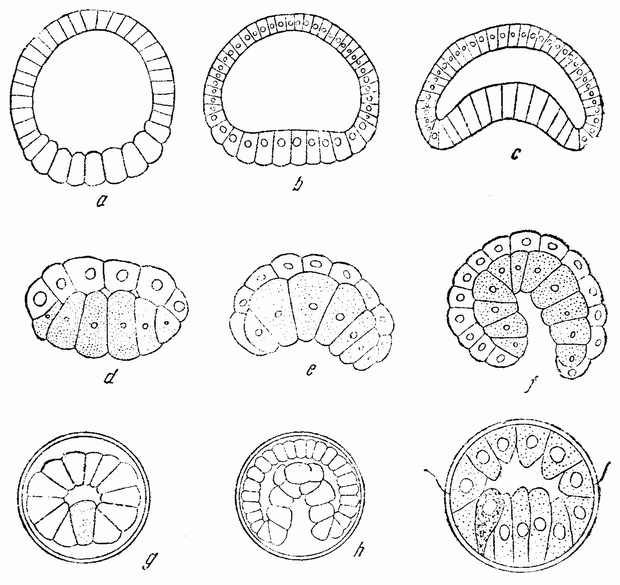 |
|
Рис. 33. а, b, с — гаструляция у ланцетника; d, e, f — гаструляция у Clavelina; g, h — гаструляция у Lucifer: i — гаструляция у Eupomatus (по Румблеру). |
щую форму чаши. С окончанием инвагинации исчезает полость бластулы; отверстие гаструлы в дальнейшем редуцируется до небольшой поры на заднем конце, которая становится анальным отверстием. Дорзальная стенка первичного кишечника ущемляется, образуя спинную струну (рис. 34, А—В). С каждой стороны спинной струны {65} образуется ряд карманов. Эти выпячивания, отшнуровывающиеся из первичного кишечника, становятся полостью тела взрослого животного (рис. 34, С) и из их внутренних стенок образуются мускульные сомиты, составляющие основную массу взрослого животного.
У лягушек и саламандр стадия бластулы (рис. 35, А) имеет над анимальным голушарием тонкую стенку, а внутри большую сегментационную полость, наполненную альбуминозной жидкостью. Нижнее полушарие состоит из крупных клеток, содержащих желток. Инвагинация начинается с одной стороны по близости от экватора, где
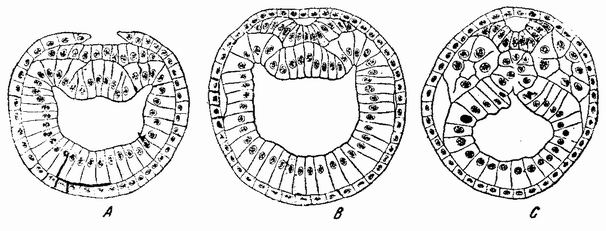 |
|
Рис. 34. Поперечные разрезы через зародыш ланцетника, показывающие образование нейральной пластинки и трубки, спинной струны и кишечных мешков (по Церфонтену); |
располагается серый полумесяц (рис. 35, D). В этом случае инвагинация по своей форме скорее подобна щели, а не открытому рту, и происходит путем втягивания внутрь клеток наружной стенки в этой области. Поверхностные клетки спереди и с боков щели двигаются к ней и вворачиваются через ее край внутрь. Опыты маркировки внешних клеток краской (рис. 36) показывают, что обширный участок наружной поверхности перемещается внутрь, закручиваясь вокруг краев отверстия, и образует верхнюю и боковую стенки первичного кишечника. Край бластопора, имеющий форму полумесяца, движется по поверхности в нижнее полушарие (рис. 37) до вегетативного полюса, или еще дальше. Первоначальная внешняя стенка этого полушария становится дном первичного кишечника, а инвагинировавшиеся клетки становятся крышей. Во время этих перемещений происходят большие смещения клеток, особенно в верхнем полушарии. {66}
Из клеток, лежащих вдоль среднедорзальной стенки первичного кишечника, развивается спинная струна, а из клеток по обеим сторонам ее развивается мезодерма или средний слой, дающий начало мускульным сомитам, полости тела и т. п. И то, и другое образуется в основном тем же путем, что и у ланцетника (рис. 34), за исключением
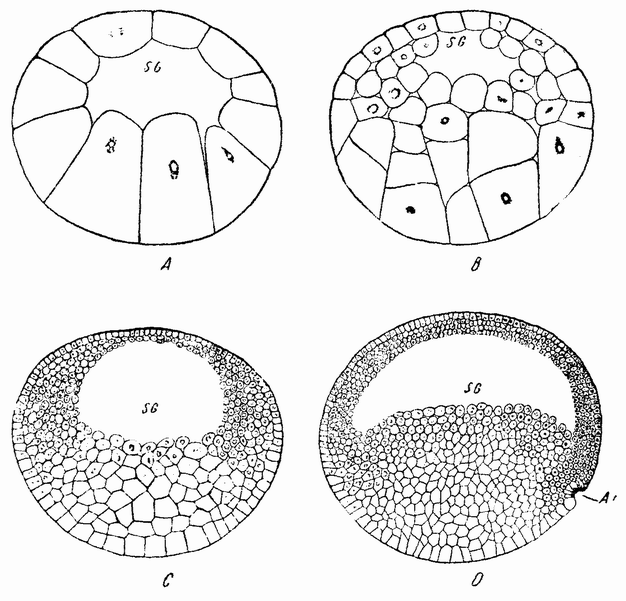 |
|
Рис. 35. Разрезы через поздние стадии дробления и начало гаструляции яйца лягушки. |
лишь того, что в этом случае не происходит образования мешков. Рот образуется в той точке, где передний конец первичного кишечника приходит в соприкосновение с наружным слоем, который вворачивается внутрь и образует внутреннюю поверхность рта. Заднее отверстие первичного кишечника или отверстия гаструлы становится анусом. {67}
У позвоночных центральная нервная система ясно выражена с самого начала развития. Она появляется у эмбриона сейчас же после окончания процесса гаструляции. Слой поверхностных клеток, лежащих впереди бластопора, начинает сдвигаться вместе, образуя удлиненной формы нейральную пластинку. Края и передний конец пластинки
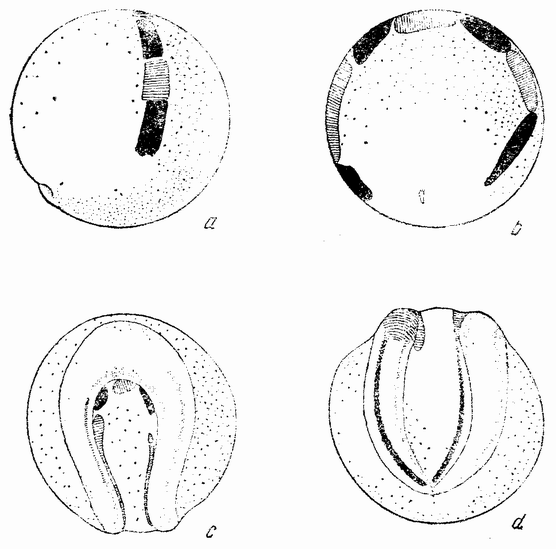 |
|
Рис. 36. а, b — стадии молодой гаструлы у тритона, на которую предварительно с помощью двух красок было нанесено полукольцо, охватывающее почти половину окружности яйца; с, d — последующее положение этих отмеченных участков (по Гертлеру). |
начинают приподыматься (рис. 36) и закручиваться, образуя полую трубку. Стенки этой трубки образуют мозг и нервный ствол, из которого вырастают нервные волокна ко всем частям тела. Нос и части органов зрения и слуха образуются инвагинацией поверхностного слоя клеток в головном участке. {68}
Зародыш птицы развивается из диска клеток на анимальном полюсе яйца, который со временем выталкивается над большой желточной массой. Первые следы гаструляции появляются на задней поверхности диска (рис. 38, а—b). Затем валик клеток, называемый первичной полоской, обозначает место бластопора (рис. 38, с), но прежде чем появляется первичная полоска, под диск углубляется пластинка клеток, дающая начало энтодерме (рис. 38, b, d).
У млекопитающих гаструляция происходит, повидимому, совсем таким же образом, как и у цыпленка, несмотря на
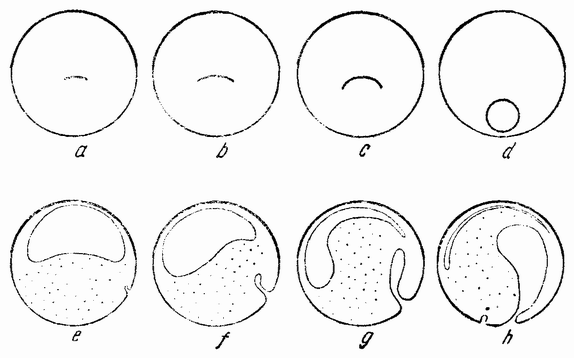 |
|
Рис. 37. Гаструляция у лягушки или тритона, а—d — вырост дорзальной губы бластопора; е—h медиальный разрез через те же стадии. |
то, что яйцо млекопитающего мельче и не содержит желтка. И все же оно развивается так, как будто бы в нем, как у ящериц и птиц, присутствует большое количество желтка. Диск клеток, из которого развивается эмбрион, лежит на поверхности большого пузырька (рис. 29, а). Периферические клетки диска простираются вокруг внутренней стенки пузырька (рис. 29, b), как будто бы для того, чтобы охватить, в действительности не существующую, большую массу желтка. Тем временем на бластодерме появляется первичная полоска (рис. 29, b) и из ее переднего конца происходит врастание клеток внутрь для образования спинной струны. Первичный кишечник и мезодерма образуются из нижнего слоя диска совсем таким же образом, как и у птиц. Процесс {69} гаструляции у млекопитающих дает много указаний на то, что он протекает по тому же пути, что у ящериц и птиц, но в то же время он выглядит как бы несколько сокращенным по сравнению с гаструляцией у последних. Не зная развития этих позвоночных, млекопитающие могут показаться одиноко стоящими в ряду остальных позвоночных,
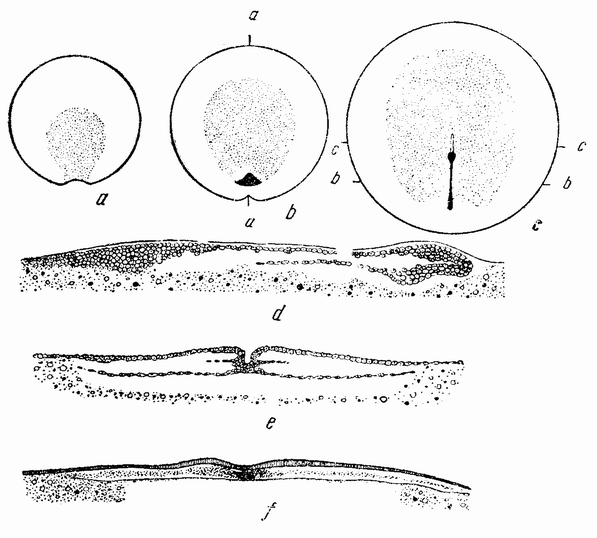 |
|
Рис. 38. а, b, с — стадии развития бластодермы цыпленка; а — продольный разрез через b; поперечный разрез через первичную борозду с; f — поперечный разрез через спинную струну с (по Пэтену). |
зная же его, мы можем объяснить происходящие изменения предположением, что хотя зародыш млекопитающего и потерял свой желток, он все же сохранил тот способ гаструляции, которому следовали его предки. Обрастание несуществующей желточной массы для образования «желточного мешка» (рис. 31, b) происходит совсем так же, как у птиц (рис. 31, a). В конце концов этот мешок, как и у птиц, втягивается в первичный кишечник. Это является {70} характерным примером сохранения процедуры, свойственной предкам, но сейчас бесполезной — желточный мешок является рудиментарным органом.
Существовали многочисленные попытки найти механическое объяснение не только гаструляции, но и вообще явлению инвагинации, играющему важную роль в эмбриональном развитии. Эти объяснения основывались почти исключительно на аналогиях с механическими моделями и очень мало на результатах опытов с живыми яйцами. Однако основой для большинства гипотез послужил следующий подмеченный факт. Когда слой цилиндрических клеток инвагинируется для образования чаши, внутренние концы клеток становятся шире, а наружные уже (рис. 33, с). Другими словами, отдельные клетки принимают форму усеченного клина. Итак, если в одном участке эластичной наружной стенки бластулы клетки так изменят свою форму, что станут клинообразными, то произойдет автоматическое впячивание стенки внутрь. С другой стороны, если стенка была вдавлена внутрь снаружи или втянута изнутри, то клетки должны принять такую же самую форму, так как внутренняя поверхность становится больше наружной. Наблюдаемые изменения формы клеток будут в обоих случаях одинаковыми. Имеются, однако, некоторые соображения, которые, повидимому, исключают последнюю гипотезу. Так, например, не известно внешних сил, на которые можно было бы сослаться для объяснения вдавливания внутрь более толстой части стенки. Если бы такое воздействие имело место, то опустившимися клетками должны были бы быть более мелкие клетки крыши гаструлы. Если жидкость полости бластулы отсасывается, выдавливается наружу, или абсорбируется стенками, то опять-таки в результате этого опустится скорее крыша, а не более толстое основание бластулы.
Противоположная гипотеза предполагает, что более крупные клетки бластулы становятся шире либо потому, что они абсорбируют воду из полости бластулы, либо же потому, что на внутренней поверхности бластулы уменьшается поверхностное натяжение. С первого взгляда нельзя объяснить, почему набухание ограничено внутренней стороной клеток, т. е. почему не набухает вся клетка и не становится сферической. Конечно, внутренняя и наружные поверхности, вероятно, отличаются друг от друга. Казалось бы, что скорее внутренняя, а не наружная, часть должна {71} абсорбировать воду, но неясно, почему такое изменение должно иметь место не раньше, а именно в этот специфический период.
Конструировались различного рода модели, которые претерпевали инвагинацию, когда создавались такие условия, что в одной части стенки внутренняя поверхность ее абсорбирует жидкость изнутри скорее, чем наружная поверхность (рис. 39). В этих условиях происходит вворачи-
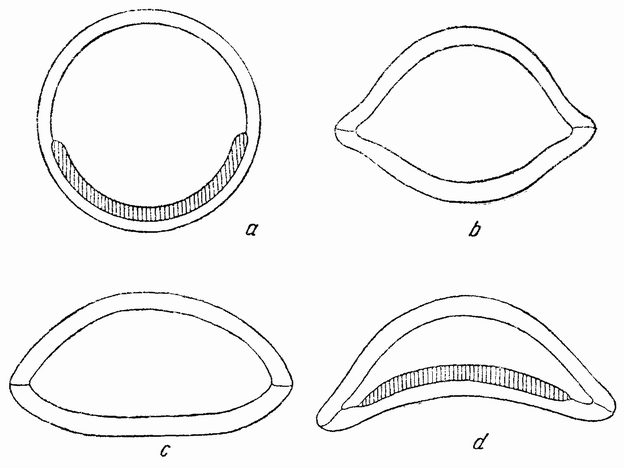 |
|
Рис. 39. Модель из желатиновых полушарий для имитации гаструляции (по Шпеку). |
вание, но это еще не доказывает, что такого же рода различия существуют и между внутренней и наружной поверхностями стенки бластулы.
Пожалуй, наиболее правдоподобная гипотеза заключается в том, что инвагинация является результатом различий в поверхностном натяжении на внутренней поверхности стенки бластулы, обусловленных скоплением внутри углекислоты или каких-то других веществ. Как результат понижения поверхностного натяжения будет происходить перемещение клеточных веществ в ту сторону, где это {72} понижение происходит, причем внутренние концы клеток будут расширяться и инвагинация последует механически. Ни одна из этих гипотез в настоящее время не является удовлетворительной, потому что отстутствуют сведения относительно природы этих клеток; но они по крайней мере достаточны для того, чтобы считать, что Процесс гаструляции не исключает возможности физического объяснения. Как и в столь многих других случаях, имеющих место при развитии, анализ явления сводит его к поведению отдельных клеток; однако наблюдаемый факт, что активность всех клеток данного участка направлена к общей цели, побуждает нас искать такое объяснение этому явлению, которое трактует начало изменения как реакцию в ответ на внешние для клетки воздействия, а не как какие-то изменения, индивидуально происходящие в клетках. Но последняя альтернатива, однако, не была бы исключена, если бы удалось показать, что в этот определенный момент развития клетки данного участка индивидуально претерпевают внутреннее изменение, делающее их соответствующими физическим условиям системы в это время. Эти изменения, можно думать, являются кульминационной точкой тех изменений в протоплазме, которые начинаются в момент оплодотворения, или же они могут рассматриваться как новые изменения, которые вызываются в это время генами.
| << | {73} | >> |
В классических опытах Вильгельма Ру (1883), вокруг которых в течение многих лет происходила большая дискуссия, один из первых двух бластомеров яйца лягушки повреждался раскаленной иглой. Другой же бластомер развивался и образовывал половину эмбриона. Ру сделал из своих опытов вполне естественный вывод, что части зародыша закладываются, начиная уже с первого деления, путем диференциальных дроблений. Он пошел даже дальше и приписал источник этой диференциации качественному распределению хроматинового вещества. Позднее Вейсман на этом же самом допущении основал всю свою теорию процесса развития. Однако даже в те времена имелось уже много данных в пользу того, что, как при первых, так и при всех последующих делениях яйца, каждая хромосома продольно расщепляется на две в точности равные части. Это последнее заключение полностью подтвердилось открытыми впоследствии генетическими фактами, которые по существу доказывают не только видимое деление хромосом, но также невидимое деление самих генов.
Несколькими годами позже Ганс Дриш (1891) экспериментировал с яйцами морского ежа и обнаружил, что если разделить первые две клетки, то каждая из них развивается в целый эмбрион половинной величины. Этот результат находился в резком противоречии с полученным Ру и приводил к противоположному выводу, а именно, что при первых делениях яйца диференцировка не имеет места. По выражению Дриша, каждая из первых двух клеток является «тотипотентной». Однако эти два опыта Дриша и Ру были неидентичными. В яйце лягушки поврежденный бластомер, остававшийся еще живым, соприкасался со своим партнером, тогда как в яйце морского ежа два {74} бластомера полностью разделялись. Это различие, однако, не должно сильно влиять на сделанный Ру вывод, так как, если диференцировка начинается с первого деления, то это должно относиться к обоим случаям.
Со времени этих первых опытов и их интерпретации было проведено много других сходных экспериментов не только с яйцами лягушки и морского ежа, но и с яйцами многих других видов. Прежде чем перейти к обсуждению относительной значимости споров, которые были основаны на этих наблюдениях Ру и Дриша, необходимо рассмотреть последние опыты.
Когда первые два бластомера морского ежа отделяются друг от друга, каждый из них округляется. При следующем делении каждый бластомер делится точно так же, как он
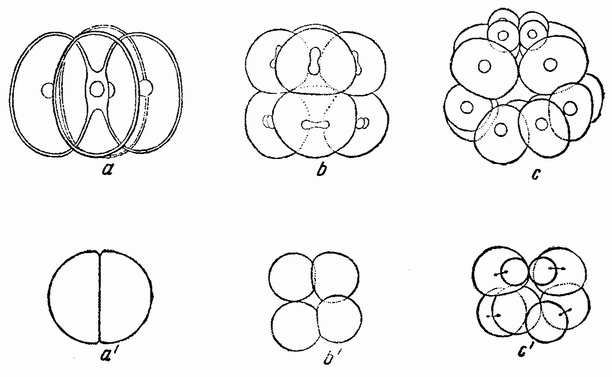 |
|
Рис. 40. а, b, с — четырех-, восьми- и шестнадцатиклеточная стадии яйца морского ежа; а1, b1, с1 — первые три дробления изолированной половины бластомера (по Дришу). |
делился бы, если бы он оставался соединенным со своим соседом, т.е. борозда проходит через анимальный полюс (рис.40, a1). Следующее деление является экваториальньш, а следующее за ним образует у вегетативного полюса микромеры (рис.40, b1—с1). При первых делениях открытая сторона {75} половинной бластулы закрывается и, по мере того как продолжаются деления, внутри бластулы появляется пространство, заполненное жидкостью. Затем происходит га-
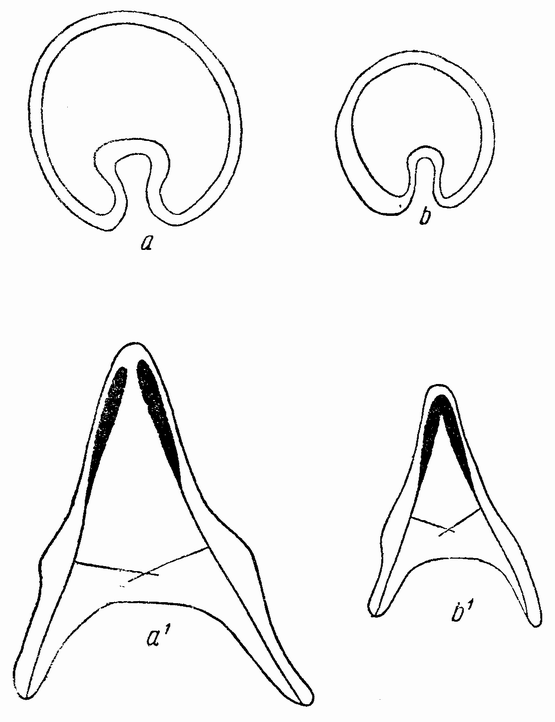 |
|
Рис. 41. Целые и половинные гаструлы и плутеусы морского ежа; последние развились из половинных бластомеров. |
струляция, после чего образуется типичный плутеус (рис. 41, b—b1).
Способ закрытия бластулы и инвагинации первичного кишечника представляет в некоторых отношениях {76} теоретический интерес. Эти процессы могут быть изучены путем окраски вегетативной области половины шестнадцатиклеточной стадии. Полученные результаты показывают, что открытая сторона половины бластулы закрывается и клетки анимального и вегетативного полюсов (равно как и клетки с боков бластулы) сближаются. Инвагинированный первичный кишечник начинается в середине энтодермального поля. Эти отношения показаны на рис. 42, на котором изображена целая бластула (b), ее нормальный способ
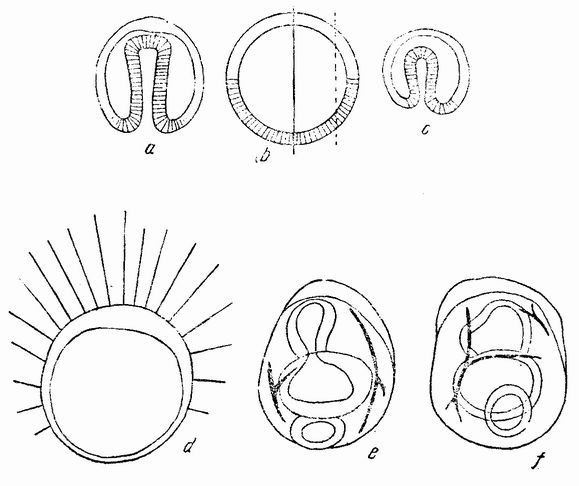 |
|
Рис. 42. b — нормальная бластула и способ ее инвагинации (а), с — для сравнения инвагинация половинной бластулы (по Герстадиусу). |
гаструляции (а) и гаструляция половинной гаструлы (с). Инвагинация происходит симметрично, причем впячивается только энтодермальное вещество. Это, казалось бы, означает, что первоначальный центр энтодермального вещества (которое после разделения лежит на стороне половинной бластулы) не является центром гаструляции половинной бластулы, а что этот центр восстанавливается в середине энтодермального вещества. Это смещение центра можно представить следующим образом. Клетки на анимальном полюсе приходят в соприкосновение с клетками {77} вегетативного полюса, где сначала был центр энтодермы. Если затем, благодаря этому соприкосновению, энтодермальные клетки теряют некоторые из своих способных к диффузии веществ, и, наоборот, эктодермальные клетки отдают энтодерме некоторые из своих веществ, то наиболее концентрированное вещество энтодермы окажется лежащим ближе к середине энтодермальной массы. Образуется новый центр, который становится центром гаструляции. Если на четырехклеточной стадии яйца морского ежа изолировано четыре бластомера, то каждый сегмент является частью целого. При втором делении образуется только единственный микромер. Развивается бластула вчетверо меньшей величины, которая гаструлируется и превращается в эмбрион-плутеус, величина которого тоже вчетверо меньше. Правда, не было обнаружено, что все четыре бластомера одного и того же яйца ведут себя одинаковым образом, но надо думать, что это так.
Надо отметить, что как половинки, так и четвертушки бластомеров содержат все вещества яйца от анимального до вегетативного полюсов, но после третьего деления четыре верхние клетки оказываются содержащими только анимальное вещество, а нижние клетки только вегетативное. В действительности было обнаружено, что некоторые из изолированных восьмушек бластомеров гаструлируют, тогда как другие остаются плавающими бластулами. Однако потенции верхних и нижних клеток лучше всего видны при отделении четырех верхних бластомеров от четырех нижних. Было найдено, что четыре верхние (анимальные) клетки дают начало плавающей бластуле (рис. 42, d), которая не гаструлирует, но у которой может произойти инвагинация рта. Четыре нижних клетки дают начало гаструле, у которой впоследствии развивается пищеварительный тракт, разделенный на характерные три отдела. Может происходить и начало образования скелета, но рот все же не образуется (рис. 42, е—f). Из этого очевидно, что верхняя и нижняя половины развиваются только в частичные структуры, сходные с теми частями, которые развиваются из этих же самых участков целой бластулы.
У асцидии положение вещей несколько отлично от такового у морского ежа, ибо, хотя у обоих дробление резко неравномерное, у асцидии гаструляция начинается на {78} более ранней стадии, причем она сначала приближается к типу, дающему половинную гаструлу, и лишь затем этот процесс до некоторой степени выравнивается.
С помощью встряхивания сегментирующихся яиц можно убить один из бластомеров, не повреждая других. В этом случае поврежденный бластомер действительно убит, а не просто устранен от развития, как это имело место в опытах с яйцами лягушки, хотя в некоторых яйцах это
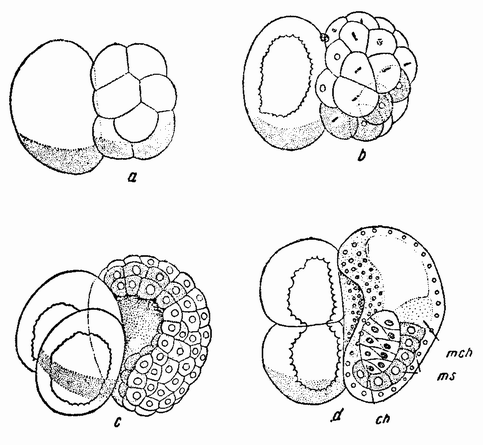 |
|
Рис. 43. Дробление, гаструляция и образование зародыша из половинного бластомера Styeia. Поврежденный бластомер не мертв, но его развитие запаздывает. Он еще соединен с более развитой половиной (по Конклину). |
все же может случиться. В одном опыте бластомеры были разделены полностью, и было показано, что они ведут себя таким же образом, как в тех случаях, когда один из них убит. Клетка за клеткой, изолированная половина бластомера претерпевает те же самые деления, которые претерпела бы эта половинка, если бы она осталась соединенной с другим живым бластомером (рис. 43). Поэтому в то время, когда зародыш начинает гаструлировать, половинка содержит как раз половину того числа клеток, которое должно быть в целом эмбрионе, и гаструляция {79} происходит, если можно так выразиться, только с одной стороны вокруг рта гаструлы, а не со всех сторон его, как это имеет место в целом эмбрионе. В тот момент, как только инвагинировались клетки первичного кишечника, спинной струны, мускулатуры и начинают сближаться поверхностные клетки, предназначенные для образования нейральной пластинки, наблюдается замечательный факт. Первичный кишечник оказывается не половинной, а цельной округлой полостью, спинная струна образует единый закругленный валик клеток, а нейральная пластинка образует единую билатерально-симметричную структуру. Другими словами, некоторые органы оказываются не половинными структурами, а вполне или почти целыми. С другой стороны, клетки, предназначенные для образования мускулатуры, не дают правых и левых структур, а образуют их только с одной или с другой стороны зародыша в зависимости от того, произошел ли эмбрион из правого или левого бластомера. Неспособность предетерминированных мускульных клеток вести себя так же, как ведут себя и другие клетки, а именно образовывать целый орган половинной величины, может быть правдоподобно объяснена их положением во время инвагинации клеток спинной струны. Мускульные клетки лежат с одной стороны спинной струны и для них может быть механически затрудненным передвигаться вокруг спинной струны для образования мускулатуры с другой стороны ее. Как бы то ни было, факт остается фактом, что дробления, подобные дроблению яиц морского ежа, находятся в строгом соответствии с той частью яйца, в которой они происходят, и эмбрион, развившийся из любой из первых двух клеток, значительно более похож на целую структуру, чем на половинную. Во многих отношениях он развивается совсем таким же образом, как и зародыш морского ежа. В обоих случаях развитие связано с сильно выраженной саморегуляцией.
Если у асцидии на четырехклеточной стадии изолированы две клетки, лежащие на дорзальной стороне яйца, на которой располагается полумесяц, то эти клетки продолжают сегментироваться, как будто бы они являются частью целого (рис. 44, а, b). Эмбрион гаструлируется, но после закрытия бластопора его развитие заканчивается. Нейральная пластинка присутствует в полной мере, но не заворачивается для образования трубки, Клетки спинной {80} струны тоже присутствуют, но не удлиняются для образования валика. Имеется и первичный кишечник. Статическое состояние этих органов может объясняться их неспособностью вытягиваться. Эмбрион не имеет мускульных клеток, каудальной энтодермы и мезенхимных клеток.
При изолировании двух других бластомеров, лежащих напротив полумесяца, они дробятся как часть целого
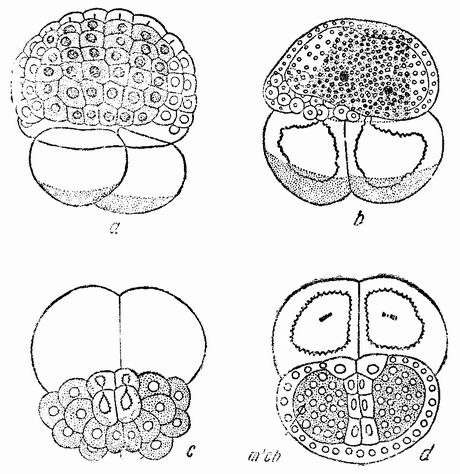 |
|
Рис. 44. а и b — развитие двух передних бластомеров Styela; с и d — развитие двух задних бластомеров; m, ch — мезенхима (по Конклину). |
(рис. 44, с—d). Мускульные клетки образуются позднее. Появляется также каудальная энтодерма и мезенхимные клетки. Следы спинной струны или нейральной пластинки отсутствуют и не происходит образования хвоста. Об этих двух эмбрионах можно сказать, что один из них потому не развивается в целый зародыш или хотя бы даже в более или менее нормальную форму, что в отсутствии другой половины он не может вытянуться, а другой потому, что он не получил в придачу от второй половины спинную струну и нейральную пластинку, как это происходит с этим участком в нормальном зародыше. {81}
Дробление одной четвертушки бластомера зависит от того, из какой части квадранта она происходит, являясь в любом случае строго неравномерным. Дальнейшее развитие ее подобно таковому у только что описанных двух соединенных бластомеров.
В свете отчетливых результатов, полученных на яйцах асцидий, могут быть лучше поняты результаты, полученные Ру на яйцах лягушек. Более того, опыты с яйцами другой амфибии — тритона, дают дальнейшие факты, касающиеся этой же самой проблемы. Плоскость первого
 |
|
Рис. 45. а — стадия гаструлы яйца тритона, которое на двухклеточной стадии было перетянуто волоском по линии будущей средней плоскости зародыша; b — подобное же яйцо, которое тоже было перетянуто на стадии двух клеток, но у которого первая плоскость дробления была фронтальной, т. е. проходила под прямым углом к средней плоскости, являющейся в яйце тритона более обычным положением плоскости первого дробления (по Шпеману). |
дробления яйца тритона обычно параллельна плоскости серого полумесяца, но средняя плоскость эмбриона соответствует не этой плоскости дробления, а плоскости борозды второго дробления, которая проходит через середину полумесяца. В некоторых яйцах тритона плоскость борозды первого дробления проходит через середину полумесяца и тогда она соответствует средней плоскости зародыша. Сначала надо рассмотреть последний случай. Если такое яйцо перевязать волоском в плоскости первого дробления таким образом, чтобы на яйце была перетяжка (рис. 45, а), и если волосок затягивать по мере дальнейшего деления яйца, то в результате разовьются два целых {82} эмбриона половинной величины. Бластопор появляется в каждой части в области половины полумесяца и, по мере того как его губы обрастают желток, вместо того чтобы следовать линии перетяжки между половинками, заворачивается в каждой части несколько в сторону от этой линии. Поверхностные клетки снаружи губ инвагинируются, как у нормальной целой гаструлы. Нейральная пластинка образуется над инвагинировавшейся хордальной энтодермой.
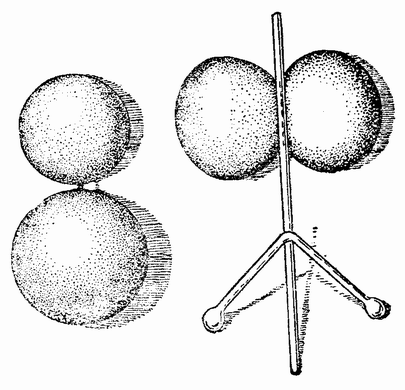 |
|
Рис. 46. Стадия двух клеток яйца тритона, оболочка которого была удалена (слева). Два бластомера разделены с помощью стеклянной палочки (справа) (по Мангольду). |
Такой же точно результат получается, если вдоль плоскости первого дробления положить стеклянную палочку (рис. 46, справа), которая с помощью грузика медленно вдавливается между двух развивающихся половинок.
Иное получается, когда первое дробление происходит в плоскости, параллельной серому полумесяцу (рис. 45, b). В этом случае в половинке, содержащей полумесяц, губа гаструлы развивается в области полумесяца и образуется целый эмбрион половинной величины. Другая же половина не гаструлирует и не образует зародыша. Из этих результатов ясно, что для того, чтобы развитие шло дальше, важно наличие области серого полумесяца или хотя бы половины его. {83}
Значение этого участка показано также другими опытами, в которых сращивались вместе половинки двух яиц, каждая из которых содержит кусочек участка губы гаструлы. Например: если два эмбриона, каждый из которых находится на стадии гаструлы, разрезаны на две части вдоль средней оси и правая часть одной половины соединена с другой половиной в соответствующем же положении (т. е. сторона к стороне), то может возникнуть единый эмбрион. Но если половинки содержат целую дорзальную губу и оси обоих соединенных половинок повернуты немного в стороны друг от друга, как показано на рис. 47, а,
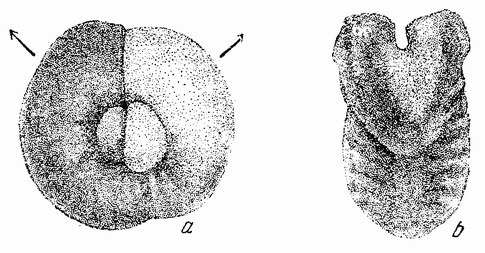 |
|
Рис. 47. а — соединение двух половинок гаструлы с дорзальными губами, повернутыми под углом друг от друга; b — зародыш с двумя головами и одним туловищем, образовавшийся в результате такого соединения (по Шпеману). |
то образуется эмбрион с двумя головами и одним туловищем (рис. 47, b).
Если у обоих гаструл отрезать верхние половинки, а нижние половинки, каждая из которых содержит целую губу гаструлы, соединить вместе так, как показано на рис. 48, а, т. е. таким образом, чтобы их оси образовывали друг с другом небольшой угол (как показано стрелками), то разовьется зародыш, имеющий одну голову и два хвоста (рис. 48, b, с). Из этого следует, что вещества, инвагинированные в месте образования губ гаструлы, продвинулись вперед и слились, образуя единую структуру, тогда как на заднем конце они остались раздельными и из каждого развилась отдельная структура.
Еще более интересная комбинация получается при соединении двух половинок гаструл, у которых передние концы каждой дорзальной губы лежат как раз друг против друга {84} (рис.49, с). В этом случае образуется эмбрион с двумя головами (рис. 49, b), каждая из которых состоит из частей каждой половины, и с двумя туловищами — по одному из каждой половинки (рис. 50, а, b). Здесь инвагинирован-
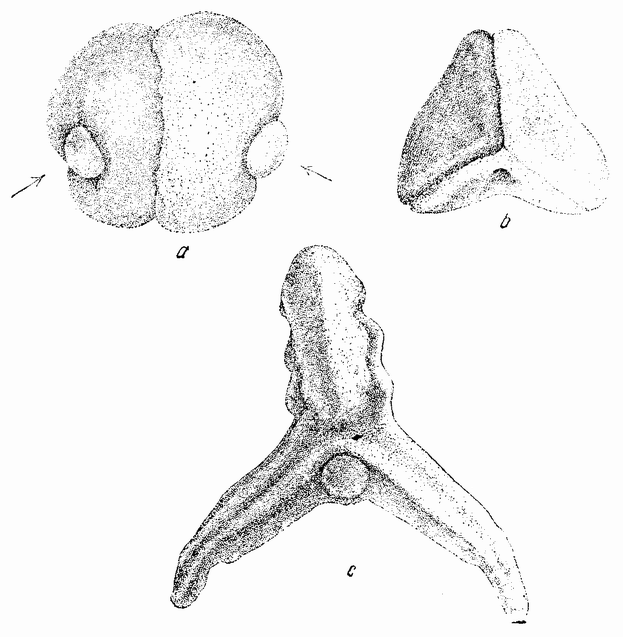 |
|
Рис. 48. a — соединение двух половинок гаструлы с дорзальными губами, повернутыми под углом друг к другу, как показано стрелками; b — стадия нейрулы, развившаяся из такого зародыша; в этом случае в результате соединения материала обеих гаструл образуется единственная голова (на рисунке каждая из соединенных половинок различно окрашена); с — более взрослая стадия того же самого зародыша с одной сложной головой и двумя независимыми туловищными отделами, по одному для каждой половины (по Шпеману). |
ные материалы, двигаясь вперед, встречаются на линии соединения и одна половина каждого вывертывается наружу для образования одной головы, а другая половина — для образования другой. {85}
Развитие яйца лягушки настолько подобно разлитию тритона, что кажется вероятным, что если один бластомер полностью удалить, то из другого разовьется полный эмбрион. Это и происходит в действительности. В большин-
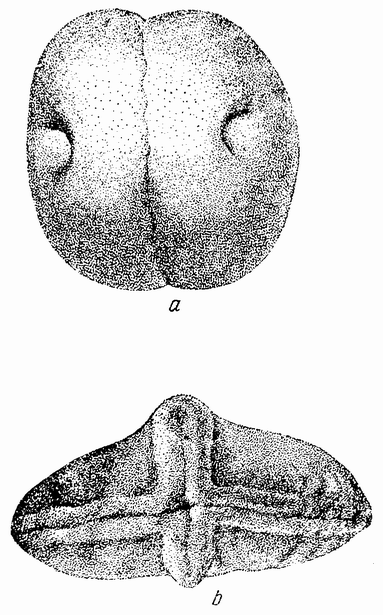 |
|
Рис. 49. а — соединение двух половинок гаструлы тритона, дорзальные губы которых направлены точно друг против друга; b — развившийся из них зародыш, имеющий две головы, каждая из которых образована из материалов обеих половинок и повернута в противоположных направлениях, и два раздельных туловищных отдела по одному из каждой половины. Срединная плоскость голов проходит под прямым углом к таковой двух туловищных участков. В результате получается типичный крестовидный зародыш (по Шпеману). |
стве яиц было невозможно отделить один бластомер, не повреждая другого, но в одном из трех опытов удалось содержимое одного из бластомеров осторожно высосать. {86}
Оставшийся бластомер развился в целый зародыш половинной величины.
Способность каждого из первых двух бластомеров яйца лягушки развиваться в целый зародыш была косвенно показана путем инвертирования яйца после наступления стадии двух клеток и удержания его в этом положении в течение последующих стадий (рис. 51, а—е). Известно, что в этих условиях вещества внутри обоих бластомеров
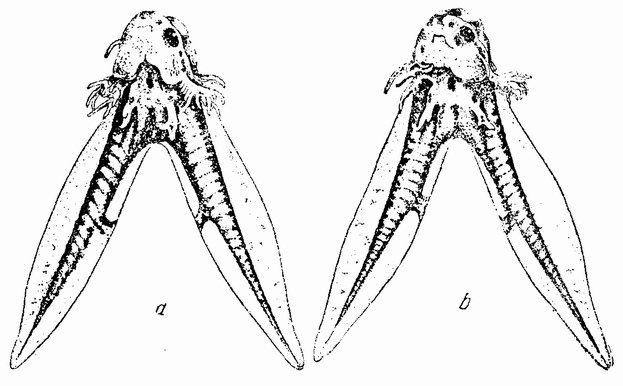 |
|
Рис. 50. Противоположные стороны крестовидных зародышей тритона, образованных в результате соединения двух половинок гаструл, как это изображено на рис. 49 (по Шпеману). |
сами перемещаются в соответствии с их весом. Более тяжелый желток при этом опускается вниз вдоль плоскости деления (рис. 51, b, с), отодвигая вещество серого полумесяца в одном бластомере от такового в другом. Каждая половина образует затем губу гаструлы и дает начало целому зародышу, развитию которого лишь несколько мешает соединение половинок (рис. 51, е). Описывалось несколько типов двойных эмбрионов, в зависимости от того, как раздвигалось вещество полумесяца (рис. 52). В свете этих данных можно теперь объяснить опыты Ру. Употребляемая им горячая игла не разрушала, а только повреждала вещества в той части яйца, в которую она вводилась. В этих условиях участок полумесяца остается {87} как целое и половина его инвапширует; поэтому вдоль линии соединения образуется половина эмбриона, как это происходило бы, если бы обе половинки развивались одновременно. Пожалуй, можно сказать, что участок серого полумесяца действует как цельный организатор, но только та часть его, которая превратилась в клетки, может продолжать развиваться в половинную структуру таким же образом, как это происходит в нормальном зародыше.
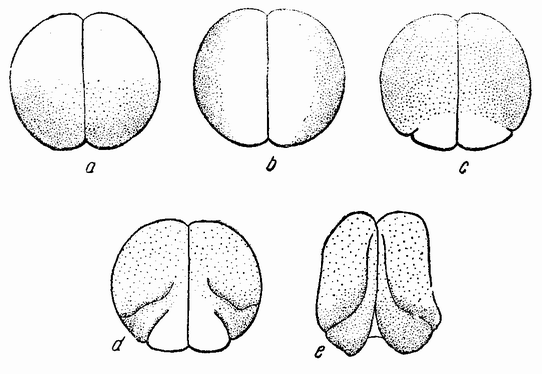 |
|
Рис. 51. Схемы инвертированной двухклеточной стадии яйиа лягушки, а — яйцо после инверзии; b — погружение желтка вдоль делящейся клеточной стенки; с — дорзальная губа, появляющаяся в каждой половине; d и е — образование двух зародышей, по одному из каждой половины. Этот способ гаструляции не находится в согласии с описанным Шлейпом процессом (по Дюркену). |
После оплодотворения в яйце ланцетника образуется плоскость билатеральной симметрии. Вещество полумесяца, которое при окраске яйца окрашивается в более темный цвет, лежит на задней стороне яйца (рис. 53, ms). Этот участок яйца становится мезодермой. Желток энтсдермальных клеток лежит впереди и вокруг вегетативного полюса (рис. 53, end). Напротив полумесяца лежит вещество, из которого образуются нейральная пластинка и спинная струна. Расположение этих веществ на четырех и восьмиклеточнои стадиях показано на рис. 53, c, d. Борозда
| {88} |
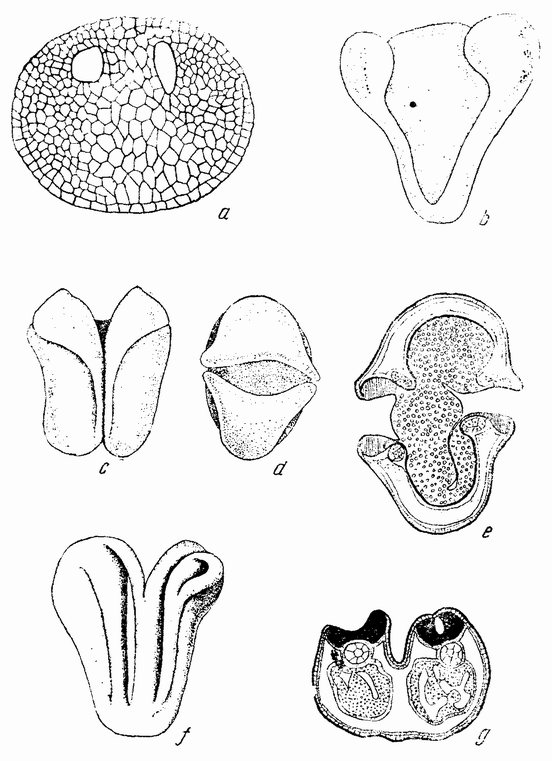 |
|
Рис. 52. Зародыши из инвертированной двухклеточнои стадии яйца лягушки, а — разрез через стадию дробления; b — два целых зародыша с противоположных сторон яйца; с — два зародыша, соединенных спина к спине; d — то же самое с переднего конца; е — поперечный разрез через последний. У этих зародышей половинные нейральные пластинки по бокам двух эмбрионов не соединились в одну нейральную трубку, как в типичном крестообразном удвоении, и они интерпретируются как два целых зародыша со spina bifida; f — два зародыша, соединившиеся боками; g — поперечный разрез через такой зародыш (по Ветцелю). |
| {89} |
первого дрооления делит яйцо на две равные части; борозда второго — на две почти равные клетки и на две меньших, лежащих с задней стороны. При третьем делении четыре клетки анимального полушария оказываются мельче, чем четыре клетки вегетативного полушария (рис 53, d).
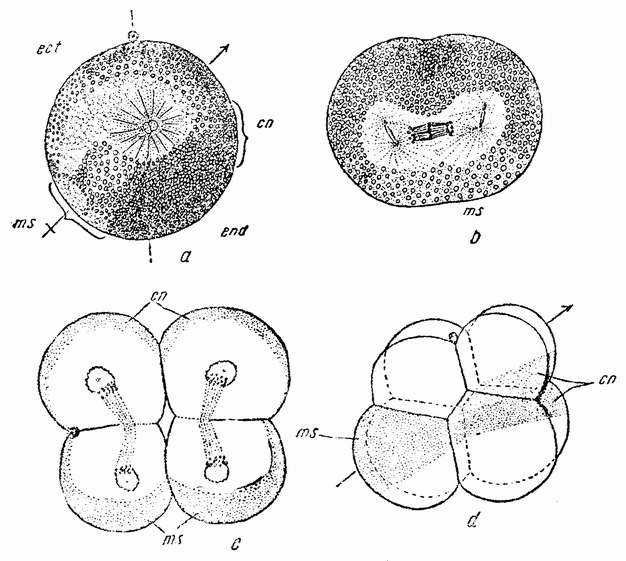 |
|
Рис. 53. Разрез через среднюю плоскость яйца ланцетника. ms — мезодермальный полумесяц на заднем конце; cn — хордо-нейральный полумесяц на переднем конце; end — эндодермальная область на дорзальной стороне; ect — эктодермальный полумесяц на вентральной стороне; b — фронтальный разрез, сделанный не задолго перед первым дроблением; с — конец второго дробления, задние клетки содержат мезодермальный полумесяц, они мельче передних клеток с хордо-нейральным полумесяцем; d — восьмиклеточная стадия, четыре микромера сдвинуты вперед над четырьмя макромерами; обозначения те же, что и раньше (по Конклину). |
Изолированные бластомеры двухклеточнои стадии, как правило, дробятся как половинки и из них развивается целый зародыш половинных размеров. Изолированные бластомеры четырехклеточной стадии тоже дают начало {90} зародышу, имеющему форму целого эмбриона, но зародыш, развивающийся из бластомеров передней четверти, оказывается частично неполным в отношении одних органов, а зародыш из задней четверти — неполным в отношении других органов. Личинка, происходящая из задней части, не имеет неиральной пластинки и хорды; личинка из передней части имеет нейральную пластинку, хорду и следы сомитов, но из нее все же никогда не образуется полная личинка. Развитие изолированных бластомеров ланцетника во многих отношениях подобно таковому у асцидий.
| << | {91} | >> |
Развитие фрагментов яиц, которые перед оплодотворением были разрезаны на части, позволяет изучать локализацию в пределах самой клетки, так же как это делалось при изолировании бластомеров. Здесь мы имеем одну из редких в биологии возможностей непосредственно изучать свойства клетки, в которой происходят регулятивные процессы организма.
Самые первые наблюдения в этой области были сделаны на фрагментах, полученных путем разлома на части яйца морского ежа сильным встряхиванием яиц в пробирке, наполовину наполненной водой. Но при этой процедуре фрагменты часто повреждаются, и так как точное происхождение каждого фрагмента остается неопределенным, то этот метод был оставлен. С помощью тонких стеклянных игол оказывается возможным под бинокуляром разрезать яйцо на две части в любом желательном месте, очень мало повреждая его. Когда разрез проходит через полюсы, его называют меридиональным, а при разрезе под прямым углом к этой оси — экваториальным.
Было сделано огромное число опытов с яйцами различных видов морских ежей, но полученные результаты не вполне единообразны. Некоторые из этих различий в результатах могут быть обусловлены различиями применявшихся методик. Так как у большинства видов полярные тельца у зрелых яиц исчезают и так как положение пронуклеуса по отношению к полярной оси изменяется, то определить плоскость разреза по отношению к оси яйца трудно или даже невозможно. Единственным методом ориентировки является воронка в окружающем яйцо студне, но наличие этого студня делает операцию трудной или неточной. Однако у одного вида, а именно у Paracentrotus lividus, в вегетативном полушарии яйца имеется пигментное кольцо {92} (рис. 14), которое делает ориентировку разреза возможной. Поэтому последующее изложение относится главным образом к опытам с яйцами этого вида.
Когда яйцо разрезано на две части по экватору, т. е. параллельно и «выше» пигментного кольца, то анимальный фрагмент (после оплодотворения) сегментируется так же, как это происходит с этой частью целого яйца, т. е. на восьмиклеточной стадии (соответствующей половине целой шестнадцатиклеточной стадии) все клетки оказываются равными. Противоположная половина, содержащая пигментную полосу, делится так же, как и целое яйцо, т. е. она обнаруживает характерные черты целой шестнадцатиклеточной стадии (рис. 54, а), а именно четыре микро-
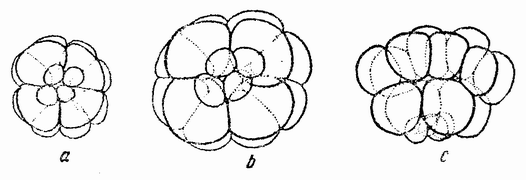 |
|
Рис. 54. Шестнадцатиклеточная стадия вегетативной половины несегментированного яйца. Схема деления такая же, как и у целого яйца на соответствующей стадии (по Герсгадиусу). |
мера (на вегетативном полюсе), четыре макромера (в середине) и восемь мезомеров (на другом конце). Другими словами, она делится не как часть целого, а как целое яйцо. Пропорциональность между рядами клеток остается при этом такой же самой, как и в нормальном яйце (рис. 54, b).
Если же яйцо разрезано на две части по оси, т. е. под прямым углом к кольцу, то дробление протекает более изменчиво. Большинство половинок образует при этом нормальную шестнадцатиклеточную конфигурацию (рис. 54, с), особенно же если яйцо было разрезано перед оплодотворением, но некоторые половинки дают различного рода неравномерные типы дробления. Разница между различными типами дробления фрагментов может быть частично обусловлена трудностью разрезать яйцо точно через полюсы, различиями в закрывании поверхности разреза, наличием на одной стороне поверхности разреза {93} и т. д. Однако основные моменты совершенно ясны. Анимальное и вегетативное полушария яйца ведут себя не одинаково, причем это различие указывает на то, что они содержат различные вещества. Благодаря этому может быть в некоторой степени понятно пропорциональное развитие в вегетативных половинках и в меньшей степени в латеральных половинках, ибо митотические фигуры, определяющие плоскость дробления, регулируются общим количеством и распределением веществ яйца, из которых они развиваются. При половинном количестве вещества в половинном фрагменте величина митотической фигуры должна быть вдвое меньшей. Эта интерпретация сама по себе недостаточна для объяснения относительной величины бластомеров, ибо микромеры, например, в вегетативной половине, могут оказаться значительно крупнее, чем это ожидается. Должны быть, следовательно, допущены дополнительные факторы. И действительно, имеются данные, что вещество, попадающее в различные бластомеры, не обособлено. Например, не все вещество, попадающее в микромеры, содержится в нормальных микромерах, а часть его простирается дальше в анимальном направлении, так как для образования микромеров оно используется не полностью. Следовательно, для того чтобы дробление было пропорциональным, должно существовать некоторое восстановление или перераспределение близких слоев перед дроблением или во время него.
Было изучено поведение фрагментов неоплодотворенного яйца асцидии — Ascydiella. В настоящее время результаты этих опытов еще недостаточно ясны, для того чтобы послужить основанием для чего-то большего, чем только самых общих положений. Когда яйцо разрезано на две части по экватору или поблизости от него, то получаются различные типы ненормальных зародышей. В анимальной половине может нехватать энтодермы, а в вегетативной половине может быть нехватка эктодермальных частей. Могут быть получены парные зародыши, из которых один состоит из эктодермы, энтодермы, мезодермы и мезенхимы, а другой из эктодермы, энтодермы, нервной системы и спинной струны.
При разрезании яйца вдоль или почти вдоль его полярной оси может получиться пара половинных эмбрионов, два симметричных зародыша или же пара эмбрионов с дополняющими друг друга дефектами. Было предположено, {94} что половинные зародыши получаются в тех случаях, когда разрезы проходят через заранее детерминированную срединную плоскость, а симметричные эмбрионы, когда разрез перпендикулярен к этой плоскости. До тех пор, пока не будут описаны более тонкие детали стадий дробления фрагментов и раннее образование органов, такая интерпретация результата этих опытов останется очень неопределенной.
У немертины Cerebratulus развитие яйцевых фрагментов было изучено более подробно, чем у какого-либо другого
 |
|
Рис. 55. Яйцо Cerebratulus во время стадий созревания, а — яйцевое ядро (зародышевый пузырек) еще присутствует, b — образовалось полярное веретено и с — выделилось первое полярное тельце (по Вильсону). |
животного. У этого объекта возможна точная локализация разреза и консистенция протоплазмы у него такова, что после операции фрагменты быстро округляются без видимых повреждений. Яйцо при откладке его самкой имеет в анимальном полушарии очень большой зародышевый пузырек (рис. 55, а), а на вегетативном полюсе удлинение студенистой оболочки и даже, в начале, короткий выступ вещества самого яйца.
Нормальное яйцо делится поровну на две, а затем на четыре клетки (рис. 56, а—b). Борозда третьего дробления образует почти равные клетки, но четыре анимальных клетки немного крупнее вегетативных (рис. 56, с). Это деление происходит спирально и против часовой стрелки. Последующие деления происходят попеременно по и против направления часовой стрелки (рис. 56, d). Позднее вегетативные клетки инвагинируют для образования гаструлы. Образуется типичная свободно плавающая личинка, называемая pilidium (рис. 57). {95}
После того как яйцевое ядро распалось для образования полярного веретена, яйцо может быть разрезано экваториально на две части и каждая часть оплодотворена. Анимальный фрагмент, содержащий яйцевой пронуклеус, выделяет два полярных тельца. Вегетативный фрагмент не выделяет ни одного полярного тельца, так как яйцевое ядро в нем отсутствует, но развивается только с пронуклеусом спермия, который служит ему «центром деления». Характер дробления обоих фрагментов во всех деталях
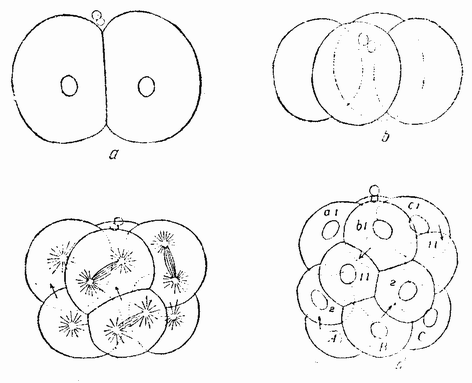 |
|
Рис. 56. Нормальное дробление Cerebratulus (по Зеленому и Вильсону). |
подобен таковому целого яйца, включая чередование спиральных делений и относительные размеры бластомеров. Тот же самый результат получается независимо от того, проходит ли экваториальный разрез у экватора или выше или ниже него (рис. 58), за исключением лишь случаев, когда анимальный или вегетативный фрагменты исключительно малы.
Результаты практически одинаковы, когда яйцо разрезано на две части, меридионально, т. е. когда разрез проходит через ось, около нее, или ближе к одной стороне. Дробления части протекают при этом нормально, но могут быть и некоторые неправильности в делениях.
Были также изучены фрагменты, отрезанные во время образования полярных веретен, когда два пронуклеуса {96} конъюгируют и после того как начала появляться борозда первого дробления. Каждый из этих фрагментов дает начало небольшому нормальному зародышу. Тем не менее кажется, что по мере приближения момента первого дробления, удаление части яйца часто ведет к отклонению от типичного пути дробления и факты неравномерного дробления становятся более и более очевидными. Этого, конечно, и следует ожидать, так как изолированные бласто-
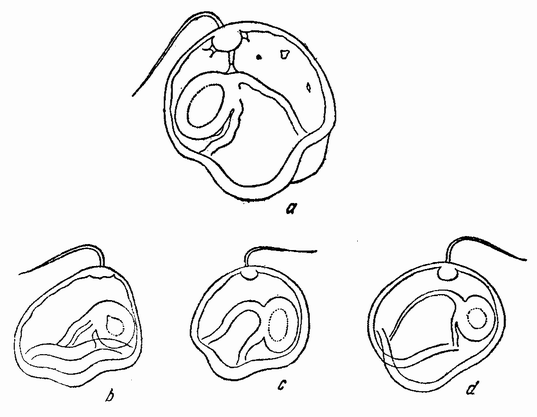 |
|
Рис. 57. а — нормальная стадия пилидиума, у Cerebratulus; b, с, d — пилидии, образовавшиеся из фрагментов яйца (по Вильсону). |
меры этих, так же как и всех других, яиц дробятся неравномерно. Эти результаты подтверждают таким образом данные, полученные при изучении других яиц, а именно, что после оплодотворения в яйце происходит прогрессивный ряд изменений, предшествующих готовому начаться дроблению. В некоторых случаях известны изменения, вызывающие видимые сдвиги веществ яйца, которые связаны с последующими явлениями. Вряд ли можно сомневаться, что сами эти перемещения зависят от уже существующего распределения веществ яйца, но в какой степени они зависят от видимой слоистости яйцевых веществ или от невидимых физических различий в различных частях яйца — в настоящее время совершенно неизвестно. Однако {97} что развитие не зависит от строгой прелокализации органообразующих материалов в яйце как целом, ясно из того факта, что часть при определенных условиях проделывает то же, что и целое яйцо.
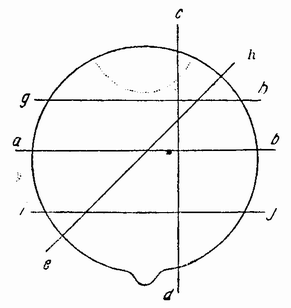 |
|
Рис. 58. Схема плоскостей разрезов через яйцо Cerebratulus (по Вильсону). |
Изучались также фрагменты неоплодотворенных яиц некоторых других животных, как-то гребневика Beroё, аннелиды Chaetopterus, моллюска Dentalium, ланцетника и тритона, но так как они не дали существенно отличных результатов от только что приведенных примеров, то их можно не перечислять.
| << | {98} | >> |
Изучение фрагментов яиц показало, что из частей одного яйца могут развиться два эмбриона. Было описано также и обратное явление — развитие единого эмбриона из двух слившихся целых яиц. Например: если сливаются два яйца круглого червя Ascaris megalocephala до образования полярных телец (рис. 59), то последние выделяются затем на анимальном полюсе каждого яйца. Если полюсы лежат близко друг от друга, то и полярные тельца располагаются рядом, но если яйца слились так, что анимальные полюса лежат раздельно, то полярные тельца выделяются тоже в различных точках. Если в такое яйцо проникает один спермин, то может случиться, что он сольется с двумя пронуклеусами и образует триплоидный эмбрион. Можно было предполагать, что такое двойное яйцо дает начало гигантскому эмбриону и такие гигантские эмбрионы, т. е. зародыши двойных размеров, были найдены. К сожалению, однако, неизвестно, развиваются ли двойные эмбрионы только, когда анимальные участки двух соединившихся яиц лежат близко друг от друга, или же они могут развиться и в тех случаях, когда они лежат далеко один от другого. Бывает также, что два яйца аскариды могут слиться вместе после того как полярные тельца уже выделились, но развивается ли при такой комбинации нормальный эмбрион — неизвестно. Наличие двух яйцевых пронуклеусов и возможность проникновения в яйцо двух спермиев вызывает усложнения, которые вероятно будут мешать нормальному развитию.
У немертины Lineus ruber описывались гигантские яйца, которые возникают в результате слияния двух яиц. Такое слияние может происходить до дробления, во время дробления или на стадии бластулы. При каких условиях слияния {99} могут развиться нормальные эмбрионы двойной величины — неизвестно.
У тритона оказалось возможным получить двойные зародыши путем соединения яиц на стадии двух клеток, принявши меры, чтобы полумесяцы яиц были совсем, или почти
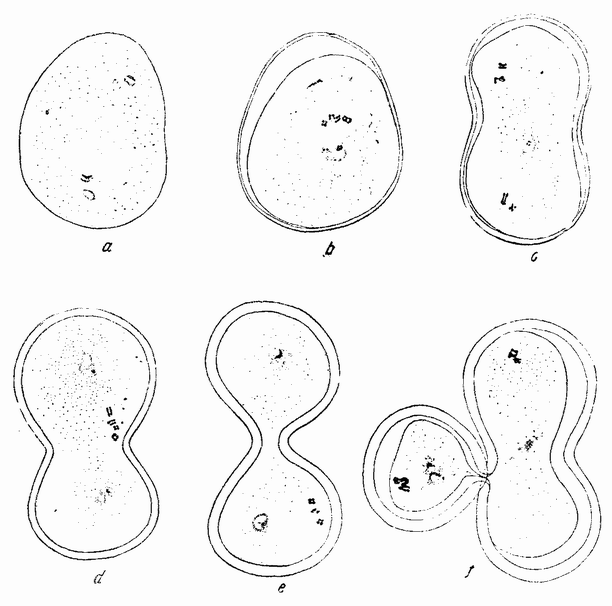 |
|
Рис. 59. Двойные яйца аскариды с полярными тельцами (диадные группы хромосом) и яйцевыми и семенными ядрами (по Коутч). |
одинаково ориентированы. Так, после удаления оболочек первые два бластомера далеко расходятся друг от друга. В это время одно яйцо накладывается поперек другого с повернутыми обоими полюсами, как показано на рис. 60 а; при приближении бластомеров друг к другу четыре клетки соединяются и такая комбинация может образовать один (рис. 60 c, d), два или даже три эмбриона. Представляется {100} вероятным, что единый эморион развивается только в тех случаях, в которых полумесяцы двух яиц оказались лежащими близко один от другого; если же этого не произошло, то могут развиться два или три соединенных зародыша. Делались многочисленные попытки получить единый зародыш морского ежа путем слияния двух яиц, либо до дробления, либо после него. В огромном большинстве случаев
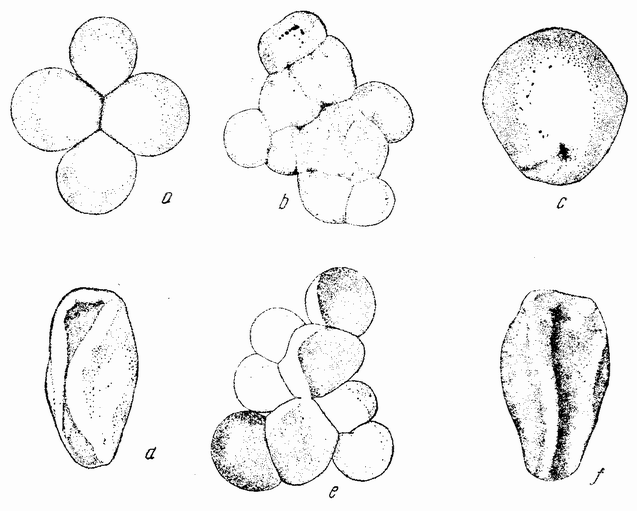 |
|
Рис. 60. а — два яйца тритона на двухклеточных стадиях, положенные поперек друг друга; b — более поздняя стадия дробления; с — единая гаструла; d — стадия нейрулы у этого зародыша; е — другая стадия дробления двух яиц, соединенных, как в a; f — стадия нейрулы (по Мангольду). |
возникающие при этом два соединенных эмбриона оказываются неполноценными в каком-нибудь одном или во многих отношениях (рис. 61 и 62). Изредка же единые эмбрионы все-таки получались (рис. 62 а). Недавно эта проблема была изучена более тщательно путем соединения двух яиц, находящихся на четырехклеточной стадии, так что полярные оси обоих яиц располагались на одной линии. {101} Ориентировка соединения обнаруживается позднее положением микромеров и инвагинацией гаструлы. В этих опытах одно из двух яиц, оболочка которого удалена, окрашивается сначала ниль-блау. Сращивание производится в лишенной кальция воде, в которой яйца имеют тенденцию прилипать
 |
|
Рис. 61. а и b — слившиеся бластулы морского ежа; с, d, е — образовавшиеся гаструлы; f—j — развившиеся из них плутеусы (по Моргану). |
друг к другу. Одно из яиц, находящееся на четырехклеточной стадии, кладется поверх другого и слегка придавливается. Три возможных при этом комбинации показаны на рис. 63 а, b, с. При соединении по схеме а, когда анимальные полюса лежат в одном и том же направлении, возникает две инвагинации, одна на вегетативном полюсе, а другая на экваторе бластулы. Более редко гаструляция в верхнем компоненте может быть подавлена, и из нижнего компонента образуется единый пищеварительный тракт. Это является {102} приближением к единому зародышу. При соединении по схеме b на противоположных концах бластулы обычно происходят две инвагинации. При схеме с может быть одна или несколько инвагинаций на экваторе либо же она мо-
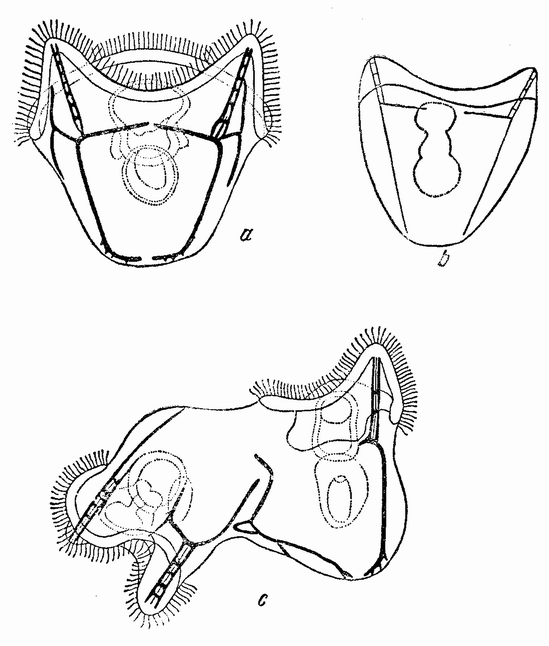 |
|
Рис. 62. Плутеусы из слившихся бластул морского ежа (по Дришу). |
жет быть подавлена в обоих компонентах. Получаются ли в некоторых соединениях более совершенные плутеусы, если слияние двух яиц происходит перед оплодотворением или немедленно после него, но до деления, чем в тех случаях, когда слияние имеет место после завершения четырехклеточной стадии, как описано выше, — в настоящее время с определенностью неизвестно. {103}
У бабочек были найдены двуядерные яйца. Каждое ядро выделяет два полярных тельца. При оплодотворении двумя сперматозоидами каждое ядро делится, они вместе дают начало ядрам зародыша и образуется единая взрослая бабочка. Эти яйца не являются вдвое более крупными и развивающиеся из них взрослы? организмы тоже не крупнее нормальных. На какой стадии происходит слияние клеток и образование единого яйца — неизвестно, но так как эти яйца имеют такую же величину, как и нормальные, то соединение должно происходить прежде чем в них скопляется желток.
 |
|
Рис. 63. Схема, показывающая ориентировку слившихся яиц по отношению к полярной оси (по Балинскому). |
В качестве редких исключений двуядерные яйца возникают также у мухи дрозофилы. Такие яйца не могут быть непосредственно наблюдаемы, но об их появлении заключают по возникновению некоторых типов мозаичных мух. Например, когда такое яйцо гетерозиготно по одной паре хромосом, то может случиться, что при выделении полярных телец одно ядро потеряет один, а другое ядро другой член гетерозиготной пары хромосом. Если два сперматозоида, оплодотворяющие такое яйцо, вскрывают наличие двух редуцированных яйцевых ядер, т. е., если они содержат только рецессивные факторы, то образуется мозаик, одна часть тела которого обнаружит один полученный от матери признак, а другая — другой. Так как мухи имеют нормальную величину, то возможно, что такие яйца возникают путем слияния на ранней стадии или, что более вероятно, в результате отсутствия деления протоплазмы ооцита после разделения его ядра. При этом сохраняется первоначальная ось клетки и это условие облегчает нормальное развитие.
| << | {104} | >> |
Если яйца морского ежа или ланцетника встряхнуть, когда они находятся на стадии двух клеток, то благодаря неполному разделению двух бластомеров, из них часто развиваются два соединенных вместе зародыша, причем каждый близнец по своим размерам оказывается вдвое меньше, чем нормальной эмбрион. Изучение стадий, через которые проходят такие близнецы, показывает, что каждый из них развивается таким же путем, как и при развитии двух изолированных бластомеров, за исключением лишь того, что их соединение в большей или меньшей степени мешает развитию. Раньше предполагалось, что все близнецы и двойные зародыши возникают в результате случайного разделения двух первых бластомеров, однако, последующие наблюдения сделали эту точку зрения для большинства случаев неправдоподобной.
Изучение развития двойных зародышей ланцетника (рис. 64) показало, что они возникают не столько в результате разделения и последующего соединения вновь двух бластомеров, сколько от смещения двух бластомеров таким образом, что их оси оказываются повернутыми в различных направлениях. Полярные отношения каждого бластомера при этом сохраняются и гаструляция происходит на вегетативном полюсе каждой половины. Если вегетативные области остаются близко друг от друга, то может развиться единый первичный кишечник с признаками раздвоенности, которая впоследствии может выправиться, но может и сохраниться. Если же вегетативные участки удалены друг от друга (как на рис. 64 a—f), то происходят две независимые инвагинации. Как результат этих разделений встречаются близнецы, соединенные всеми возможными способами (рис. 64 g—h). {105}
Близнецы могут быть получены из яиц лягушки путем инвертирования их на стадии двух клеток (рис. 51). В этом случае образование близнецов обусловлено вращением
 |
|
Рис. 64. Двойниковые эмбрионы ланцетника, а—е — гаструпяция у нескольких таких зародышей; f — тройная гаструляция; g—h — более взрослые двойниковые зародыши; g — зародыши, соединенные только в хвостовой области вентральными поверхностями друг к другу, h — зародыши, соединенные дорзальными поверхностями (по Конклину). |
содержимого бластомеров таким образом, что две половинки полумесяца оказываются разделенными и каждая впоследствии становится независимой (рис. 52 b—g). {106} Принципиально эти результаты подобны полученным у ланцетника и морского ежа, ибо не так важна полярность, как разделение областей инвагинации. У тритона близнецы могут быть получены путем перетягивания яйца на стадии двух клеток или даже еще позднее, так, чтобы область гаструляции разделялась пополам. Во всех этих случаях существенно то, что каждая отдельная часть становится новым целым вдвое меньших размеров.
В дополнение к этим случаям, в которых двойные эмбрионы были получены путем частичного разделения первых двух бластомеров, имеется другая группа случаев, где удвоение было вызвано экспериментально путем изменения направления борозды первого дробления механическим воздействием, температурой или центрифугированием. Кроме того, существенно, что удвоение в этих случаях происходит в тех яйцах, в которых после оплодотворения имеется определенная сторона, дающая начало клеткам мезобласта, играющим, повидимому, роль организаторов. У кольчатых червей Chaetopterus и Nereis и у моллюска Cumingia первое деление оказывается неравным и большая клетка содержит материал для будущего мезобласта. Если же яйца поместить перед дроблением под пресс, то во многих из них первое деление будет равным. Следовательно, мезобластная область яйца присутствует теперь в обоих бластомерах и из этих яиц развиваются двойные эмбрионы. Эти результаты весьма важны потому, что они приводят в соответствие с нормальным развитием яиц особые случаи, приводящие к удвоению. В других случаях вызывания удвоений изменением окружающей среды — например, влиянием температуры (как высокой, так и низкой), действием ультрафиолетовых лучей и т. д. — эффект изучаемых агентов до сих пор еще установлен не точно.
Пытаться применить результаты, полученные при работе с искусственной средой или другими ненормальными условиями к случаям, в которых возникновение близнецов является частью нормального развития вида, в настоящее время рискованно. Так, например, у броненосца Armadillo из единственного яйца, как правило, развиваются четыре или даже восемь зародышей. У человека тоже довольно часто встречаются случаи образования близнецов, причем тенденция к образованию близнецов у человека, повидимому, наследуется. Прежде чем перейти к {107} обсуждению этих явлений, надо остановиться на описании некоторых случаев влияний внешней среды на образование близнецов.
У костистых рыб, особенно у форелей, иногда встречаются двойные эмбрионы. Вначале они кажутся лежа-
 |
|
Рис. 65. Три бластодиска рыбы с зачатками двух эмбрионов (по Куберу). |
 |
|
Рис. 66. Двойниковые зародыши рыбы, а — соединившиеся вентрально; b — один зародыш много мельче другого (по Стоккарду). |
щими в двух различных точках по краям бластодермы (рис. 65). Если они расположены один против другого, то два возникающих зародыша оказываются впоследствии соединенными вентральными поверхностями, как это изображено на рис. 66. Если же они лежат близко друг от друга, то при нарастании бластодермы на желток, они {108} могут соединиться и образовать двухголовых близнецов с одним туловищем (рис. 67). Иногда один эмбрион может быть дефективным или меньше другого по своим размерам (рис. 66 b и 67 с). Такие двойные зародыши, несомненно, обусловлены врастанием гаструлы под бластодиск в двух отдельных точках и их последующее соединение зависит от нарастания края бластодермы. Какие внутренние или внешние факторы ответственны за стимуляцию такой двускладчатой инвагинации — неизвестно.
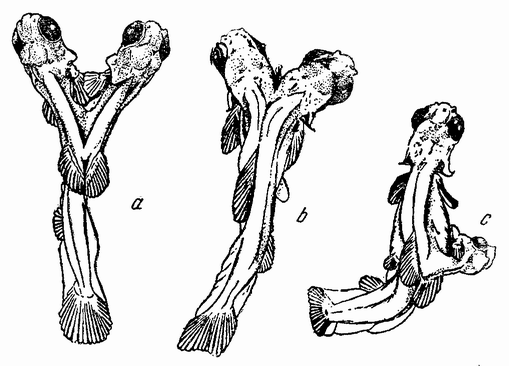 |
|
Рис. 67. Двухголовый зародыш рыбы (по Стоккарду). |
У птиц, особенно у кур, описано много случаев возникновения двойных эмбрионов и здесь имеется богатый материал для обсуждения, возникают ли они из двух бластодерм или же в двух точках одной и той же бластодермы. Возможно, что имеют место оба способа возникновения таких зародышей, но для огромного большинства случаев более вероятен последний из них. Развитие молодого эмбриона у цыпленка протекает весьма сходным с рыбами путем. Происходит раннее впячивание клеток на границе бластодермы и образование энтодермы. Затем в задней части бластодермы образуется первичная бороздка и из ее переднего конца под поверхностью растет вперед спинная струна. Поверхностные клетки спереди и вокруг первичной борозды образуют нейральную пластинку. Некоторые двойниковые зародыши у птиц имеют две головы и одно {109} туловище (рис. 68 а), одну голову и два туловища (рис. 68 b) или, что случается менее часто, они полностью разделены один от другого (рис. 68с). Нередко соприкасаться могут две головы, а два туловища могут быть втянуты в стороны в противоположных направлениях. В последнем случае образование близнецов, если это явление мо-
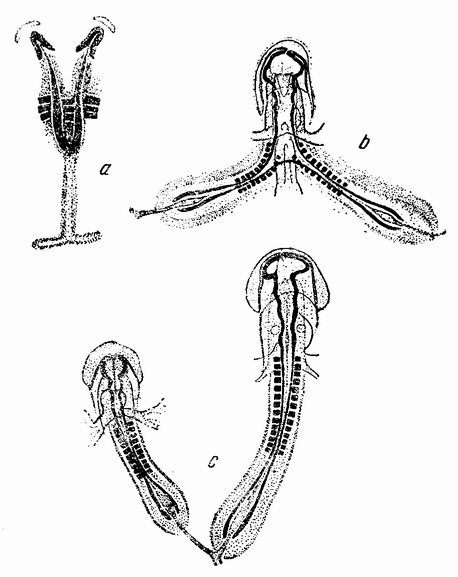 |
|
Рис. 68. Двойниковый эмбрион птицы, а — двойная голова, соединенная с туловищем; b — одна голова с двойным туловищем; с — два целых зародыша в одном яйце (по Танрейтеру). |
жет быть так названо, обусловлено, повидимому, двумя независимыми зачатками, но причина возникновения двух центров развития неизвестна. Следует отметить, что, когда разделены головной или хвостовой концы, то каждый из них имеет правую и левую сторону, но при соединении их для образования единой структуры образуется только одна правая и одна левая сторона. Другими словами, {110} аксиальные органы, нервная струна и спинная струна образуют единый медиальный орган и получается только одна правая и одна левая сторона, состоящая из участков мезобластов и других право-левых органов. Это явление весьма сходно с тем, которое встречается у описанных в главе VI двойниковых эмбрионов, образованных соединением двух половинок гаструлы яиц тритона.
Мелкие яйца млекопитающих (рис. 28) полностью делятся на две, четыре, восемь и т. д. клеток. В этом отношении их дробление более похоже на дробление ланцетника или морского ежа и можно предполагать, что разделение или сдвиг первых двух или первых четырех клеток объясняет происхождение двух эмбрионов из одного и того же яйца. Но полученные до сих пор данные не подтверждают этого взгляда, так как на ранних стадиях бластодермы не происходит доступного для наблюдения удвоения.
У Armadillo все четыре зародыша ориентированы одинаково. Четверни у этой формы (рис. 69) происходят, как известно, из единственного яйца. Надо напомнить, что хотя дробление яиц млекопитающих является полным, тем не менее последующее развитие зародыша у них очень сходно с развитием цыпленка. Поэтому можно предвидеть, что образование близнецов у млекопитающих и у птиц будет происходить сходным образом и это может быть верно и для некоторых из описанных соединенных друг с другом близнецовых эмбрионов. В наилучше изученном случае с Armadillo, у которого из одного яйца образуются четыре отдельных эмбриона, отсутствуют неправильности в расположении, подобные наблюдавшимся у нормальных близнецов птиц, а налицо закономерный процесс, приводящий к образованию четырех полных и раздельных зародышей.
У броненосца начальное развитие подобно развитию других млекопитающих. В одной точке стенки бластоцисты образуется единый клеточный шар (рис. 69 a), в котором появляется полость (рис. 69 b) и на котором впоследствии в четырех точках на равных расстояниях друг от друга вырастают четыре мешкообразных кармана (рис. 69 d). Из каждого такого кармана развивается по одному эмбриону, которые лежат по четырем меридианам большого бластоциста (рис. 69 с) и имеют общую плаценту (рис. 70). Все четыре эмбриона всегда оказываются одного и того же пола, как это и следует ожидать при оплодотворении одного яйца одним спермием. Хромосомный механизм {111} определения пола млекопитающих (XX—XY) объясняет, почему все четыре эмбриона Доказываются или самцами или самками. Было доказано, что у Armadillo яйца действительно освобождаются из яичника только по одному.
Термин близнецы в применении к человеку означает одновременное рождение двух младенцев. Большинство близнецов получаются из двух отдельных яиц, каждое из которых оплодотворено различным сперматозоидом. По-
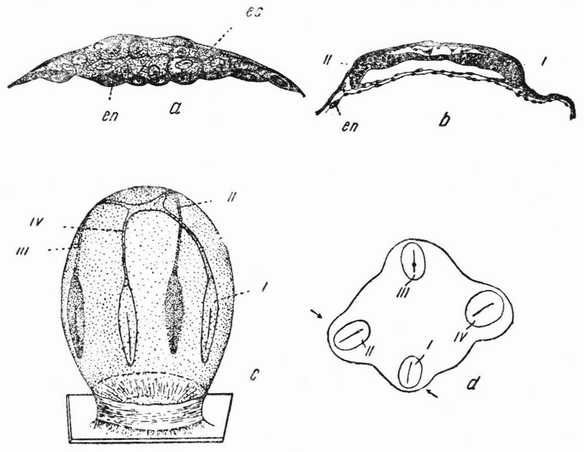 |
|
Рис. 69. а — разрез через раннюю бластодерму броненосца Armadillo; b — более поздняя стадия с образовавшимся амнионом; I и II — образование боковых карманов, в каждом из которых разовьется один зародыш; с — более поздняя стадия с четырьмя эмбрионами; d — общий вид, показывающий положение четырех зародышей (по Патерсону). |
этому такие близнецы могут быть двумя мальчиками, двумя девочками или девочкой и мальчиком. Такие близнецы называются двуяйцевыми, или дизиготическими. В других случаях близнецы происходят из одного яйца, оплодотворенного одним спермием, и тогда они называются однояйцевыми, монозиготическими, или идентичными близнецами. Последние всегда одного и того же пола, т. е. либо оба мальчики, либо оба девочки. Так как две таких особи имеют совершенно одинаковые гены, то они должны быть {112} очень сходными или даже идентичными не только но своим структурным и физиологическим признакам, но даже по своим психическим особенностям, за исключением лишь признаков, полученных ими под влиянием внешних условий. Из осторожности следует также оговориться, что какие-либо неправильности в положении двух эмбрионов по отношению к плаценте или друг к другу могут вызвать более быстрое и лучшее развитие одного эмбриона по сравнению с другим, причем эти различия в эмбриональной
 |
|
Рис. 70. Девятиполосый броненосец (Armadillo). Четыре идентичных близнеца с общей плацентой (по Ньюмаму и Патерсону). |
внешней среде могут дать одному из близнецов небольшие преимущества в начале его развития. При таких изначальных различиях последующая судьба даже идентичных близнецов может быть несколько различной.
Раннее эмбриональное развитие идентичных близнецов у человека не изучено. Однако же развитие человеческого эмбриона подобно развитию всех других млекопитающих, поэтому представляется не невероятным, что человеческие близнецы возникают довольно сходным образом с четвернями у броненосца. У обоих имеется некоторое проявление зеркальной симметрии, которая доказывает, что близнецы возникают из одного общего пузырька и являются зеркальным отражением друг друга. В связи с этим {113} следует отметить, что идентичные близнецы не всегда оказываются разделенными при рождении. Известно много случаев соединения близнецов всеми возможными способами (рис.71). Большинство таких близнецов погибает при рождении, но некоторые из них остаются жить. Классическим примером близнецов этого рода являются Сиамские близнецы.
Идентичные близнецы обнаруживают сходство в структуре до мельчайших деталей, включая даже такие признаки, как отпечатки пальцев. Это позволяет судить о силе и зна-
 |
|
Рис. 71. Человеческие близнецы, соединенные с рождения (по Вильдеру). |
чении наследственных единиц. Было бы трудно придумать лучший опыт, чем этот, для того чтобы установить влияние генов на соматические признаки. Правда, оба эмбриона развиваются при идентичных внешних условиях, но это же имеет место и у двуяйцовых близнецов, которые развиваются из отдельных яиц, оплодотворенных различными спермиями; несмотря на это, последние похожи друг на друга ге больше, чем другие дети тех же родителей, рожденные в разное время.
Для изучения человеческой психики существование идентичных близнецов открывает очень благоприятную возможность для выяснения вопроса, являются ли их психические особенности в начале развития тоже идентичными или же законы наследственности в этой области не применимы. Имеется все увеличивающееся число зарегистрированных {114} случаев, в которых идентичные близнецы с момента рождения воспитывались раздельно. Их структурное сходство оставалось при этом почти таким же, как и у воспитываемых вместе. Что же касается их психического сходства и различий, то в настоящее время нельзя сделать определенного вывода; разве только можно рискнуть сказать, что в целом среда играет менее важную роль, чем наследственность, — вывод, который Гальтон сделал еще давно в результает своих работ с близнецами. Однако вряд ли можно сомневаться, что с усовершенствованием тестов психологического поведения, которое позволит нам лучше, чем это делается сейчас, различать прирожденные способности от результатов внешнего влияния общественных отношений и воспитания, перспектива определения, что именно обусловлено прирожденными свойствами, а что приобретено извне, станет очень многообещающей.
| << | {115} | >> |
Первые исследования по экспериментальной эмбриологии привели к заключению, что для нормального развития необходим по крайней мере один набор хромосом, т. е. по одной хромосоме каждого сорта, но, как было уже установлено выше, этот вывод оставляет открытым вопрос, в какой степени будут влиять на развитие добавочные хромосомы. Что для развития достаточно только одного набора хромосом, было показано развитием нормальных эмбрионов из безъядерных фрагментов яиц, которые были оплодотворены одним сперматозоидом. Эти данные экспериментально доказывают известный уже для некоторых насекомых факт, что самцы коловраток, пчел и некоторых видов тлей начинают свое развитие с половинным числом (гаплоидным набором) хромосом. В последних случаях хромосомы происходят только из яйцевого ядра, которог развивается партеногенетически, т. е. без оплодотворения. Генетические исследования последних лет доставили много сведений об эффекте прибавления к нормальному числу хромосом одной или большего числа целых хромосом или частей их, а также показали, что получается, когда у различных видов основные (гаплоидные) числа хромосом повторены три раза (триплоиды), четыре раза (тетраплоиды) или много раз (полиплоиды).
Возникает три основных вопроса. Во-первых, об отношении изменения числа хромосом к общему количеству протоплазмы, развивающейся при этих условиях; во-вторых, о равновесии между генами и, в-третьих, что происходит во время процесса созревания половых клеток, когда сходные хромосомы конъюгируют друг с другом.
Иногда случается, что два семенных ядра из двух сперматозоидов, одновременно проникнувших в яйцо, сливаются с яйцевым ядром, в результате чего образуется три {116} набора хромосом. Но так как каждое ядро спермия привносит с собой в яйцо по центриоли, которые делясь, образуют четыре полюса для веретен (рис. 2), то развивается сложная митотическая фигура и распределение хромосом оказывается неправильным. Однако эти случаи являются плохим материалом для анализа настоящей проблемы. Значительно более благоприятные случаи были найдены у растений, у которых пыльцевое зерно, могущее изредка содержать в себе два набора хромосом, оплодотворяет яйцо с одним набором их, или наоборот. Трудностей с центриолями у растений не возникает, и образуется триплоидная особь. Менее часто встречаются случаи, когда и яйцо, и пыльцевое зерно имеют по двойному набору хромосом, образуя особь, являющуюся тетраплоидом.
Если яйцо животного оплодотворено диплоидным сперматозоидом, то оно будет делиться обычное число раз. Если имеется какое-либо влияние хромосом на объем протоплазмы, то не ожидается, что оно проявится на этих стадиях развития. Но со временем это влияние сказывается, и клетки такой особи становятся крупнее клеток нормального диплоидного организма. Подобным же образом было в нескольких случаях показано, что у тетраплоидов отдельные клетки взрослого организма в два раза превышают объем клеток нормальной особи. Очевидно, добавочные хромосомы влияют со времс нем на количество образующейся протоплазмы.
По своим признакам тетраплоидная особь в основном подобна диплоидной, т. е. обычной особи, из которой тетраплоид возник, но в некоторых отношениях, например, по своим размерам или по соотношению частей, тетраплоидная особь может отличаться от диплоидной. Некоторых различий следует ожидать, исходя из взаимоотношений объема и поверхности. Удвоение объема увеличивает поверхность только в 1,59 раза. Одинаково ли общее число клеток у диплоидных и тетраплоидных особей, не было точно определено, но кажется вероятным, что общее число клеток у них либо равно, либо почти равно. Однако в различных органах тела это может быть по-разному.
В других случаях, в которых удвоение числа хромосом произошло на ранней стадии яйца, или в соматической клетке, благодаря слиянию половинок клетки, приготовившейся разделиться (безразлично, будь то молодая яйцевая клетка или соматическая), протоплазма может {117} считаться тоже удвоившейся. Здесь не возникает упоминавшийся выше вопрос о взаимоотношении хромосом и протоплазмы в тех случаях, когда диплоидное яйцо, достигшее своего полного размера, внезапно удваивает или утраивает число своих хромосом.
В нескольких случаях приводились данные об обратном положении, а именно об относительной величине клеток у гаплоидной особи. Изучались величина и число
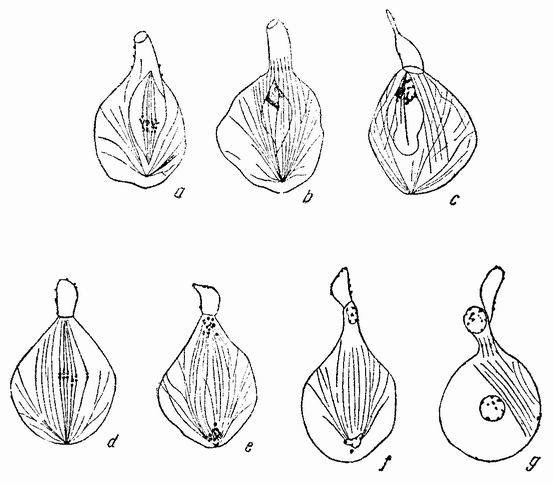 |
|
Рис. 72. Первое (а—с) и второе (d—g) деления сперматоцитов у пчелы (по Мевесу). |
клеток у самцов пчелы, являющихся гаплоидами. У взрослого самца величина клеток оказалась для каждого органа соответственно одинаковой с таковой у диплоидной самки, однако имеются некоторые данные, хотя и недостаточные для того, чтобы быть уверенным в них, показывающие, что на ранних стадиях развития самца число хромосом у него может удвоиться. Если в клетках тела самца хромосомы каким-то образом удваиваются, то в его половых клетках этого не происходит, что видно во время делений {118} созревания, во время которых присутствует резко выраженное гаплоидное число хромосом (рис. 72).
У мхов и папоротников, и в меньшей степени у семенных растений, имеется гаплоидное, или гаметофитное поколение, и диплоидное, илиспорофитное, которые чередуются в жизненном цикле (рис. 73). Обе стадии настолько различны,
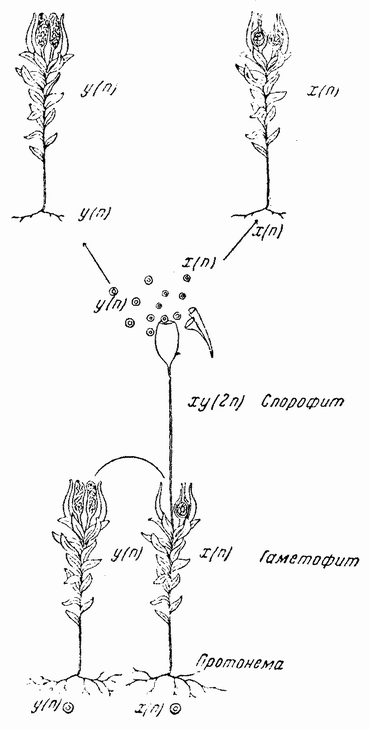 |
|
Рис. 73. Жизненный цикл мха. Мицелиальная нить и растение мха образуют гаплоидное или гаметофитное поколение; стебелек и капсула (с содержащимися в ней спорами) возникающими после оплодотворения, представляют собой диплоидное, или спорофитное поколение (по Маршалю). |
| {119} |
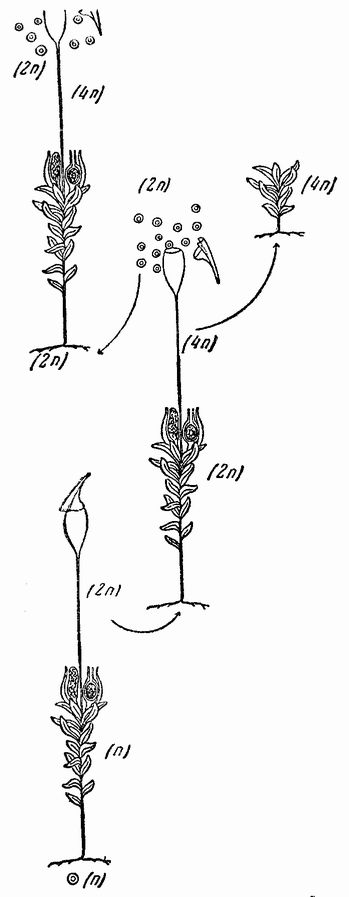 |
|
Рис. 74. Схема, показывающая образование диплоидных (2n) особей, путем регенерации спорофита у гермафродитного вида (по Маршалю). |
Последние примеры ясно показывают, что при наличии удвоенного или учетверенного набора хромосом соответственно увеличено общее количество протоплазмы. Опыты показывают также, что переход от спорофита к гаметофиту обусловливается не присутствием одного или двух наборов хромосом.
Второй вопрос касается равновесия генов. Это выражение предполагает, что общая активность генов как раз такова, что она создает характерные черты особи. Это положение может казаться тривиальностью, но истинное
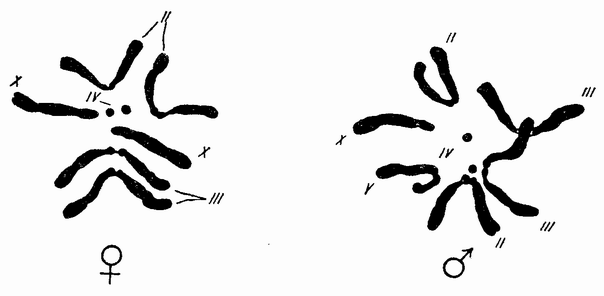 |
|
Рис. 75. Хромосомы самки и самца Drosophila melanogaster (по Добржанскому). |
значение его становится ясным, когда оно рассматривается в связи с изменениями в числе генов. Характерны два рода изменений: во-первых, все диплоидное число хромосом может быть либо умножено, либо уменьшено вдвое. Равновесие, следовательно, остается неизменным, и в этом случае можно ожидать нормального результата. Но если к полному набору хромосом добавляется одна, две или больше лишних, то равновесие изменяется. Некоторых генов становится относительно больше, чем было раньше. Примеров такого нарушения равновесия известно очень много. Иногда, например, при делении половой (или любой другой) клетки, половинки какой-нибудь хромосомы могут не разойтись к противоположным полюсам, в результате чего одна дочерняя клетка получает одну лишнюю хромосому, а в другой этой хромосомы недостает. Случись это, как это и бывает иногда, в половой клетке, то в {121} оплодотворенное яйцо будет привнесена нехватка, или дупликация хромосомы, и возникает особь, являющаяся «неуравновешенной». Такая особь может выжить и тогда она обычно обнаруживает заметные отличия от исходного типа. С другой стороны, такая особь, имеет ли она на одну хромосому больше или меньше, может и погибнуть, потому что неуравновешенное отношение генов приводит к неправильностям в ее развитии. Например, Drosophila melanogaster имеет четыре пары хромосом. Отсутствие одной из самых маленьких хромосом или прибавление третьей такой хромосомы влияет на особь так слабо, что она во всем напоминает нормальную муху, за исключением лишь немногих мелких различий. Но отсутствие одной из больших хромосом или прибавление третьей такой хромосомы влияет на развитие так сильно, что эмбрион погибает.
Явное исключение имеет место в случае половых хромосом. У некоторых форм самки имеют две Х-хромосомы, а самцы только одну и, несмотря на это, самцы и самки почти по всем их признакам очень похожи друг на друга. У дрозофилы самки имеют две Х-хромосомы, а самцы одну Х-хромосому и, кроме того, вторую половую хромосому, называемую Y-хромосомой. Было показано, что в Y-хромосоме отсутствуют почти все гены, присутствующие в X-хромосоме. Можно сказать поэтому, что в таких случаях должно было установиться некоторого рода компенсирующее равновесие, заключающееся в том, что и одна, и две Х-хромосомы дают одинаковый результат, за исключением, конечно, лишь того, что две Х-хромосомы образуют самку, а одна — самца; это различие обусловлено различием в равновесии между Х-хромосомами и остальными хромосомами.
Следующие примеры иллюстрируют зависимость пола от равновесия генов. Тетраплоидная самка дрозофилы имеет формулу 4A + 4X, а нормальная диплоидная самка — 2А + 2X. Буква А в этих формулах служит для обозначения всех остальных хромосом, кроме X. Равновесие одинаково у обоих. Тетраплоидная самка при скрещивании с нормальным самцом (2A + X) дает триплоидных самок (3A + 3X), у которых равновесие тоже такое же, как у нормальной самки. В созревающих половых клетках этой триплоиднои самки хромосомы распределяются неправильно и она поэтому полустерильна; но среди немногочисленного потомства, получающегося при скрещивании ее с {122} нормальным самцом, возникает небольшое число триплоидов, несколько больше диплоидов и немного интерсексов. Последние имеют три набора аутосом (ААА) и две Х-хромосомы (3А + 2Х). Хотя такой интерсекс имеет то же число Х-хромосом, что и нормальная самка, но других хромосом он имеет больше. Это нарушение равновесия приводит к возникновению особи в некоторой степени промежуточной между нормальными самцом и самкой. Если мы предположим, что Х-хромосома содержит больше генов, определяющих женское состояние, а другие хромосомы содержат больше генов мужских признаков, то у интерсекса равновесие оказывается промежуточным. Протоплазма яйца, являющаяся во всех этих случаях одинаковой, не играет существенной роли в этом результате.
У дурмана (Datura) оказалось возможным получить целый ряд типов, в каждом из которых имеется третья хромосома той или другой пары. Добавочная хромосома приводит к образованию у особи заметных отличий как от исходного типа, так и от каждой из других форм. Могут быть прибавлены также и две хромосомы сверх нормы, что опять-таки приводит к образованию определенного изменения.
Иногда ставится вопрос, является ли нарушение равновесия при прибавлении одной лишней хромосомы менее вредным у видов с большим числом хромосом, чем в случаях, когда хромосом меньше. Такого различия, вообще говоря, можно ожидать, но прежде чем какое-либо подобное обобщение сможет иметь большое значение, необходимо учесть индивидуальные особенности видов.
Третий вопрос касается поведения хромосом, когда они на стадиях созревания конъюгируют, т. е. соединяются в пары. При таком удвоении числа хромосом, какое имеет место у тетраплоидов, имеется по четыре хромосомы каждого сорта. Обычно они соединяются в пары, которые расходятся и делятся так же, как и при правильном созревании. Каждая из четырех получающихся клеток содержит двойное число хромосом и после оплодотворения тетраплоид повторяет себя. Такой тетраплоид является саморазмножающимся. Известно несколько случаев, в которых путем скрещивания двух видов получен гибридный тетраплоид, с одним набором хромосом одного вида и одним другого. Например, были скрещены два вида Primula, у которых каждая половая клетка имеет девять {123} (гаплоидное число) хромосом; гибрид имел восемнадцать хромосом. Это было мощное растение, но при созревании его половых клеток хромосомы, происходящие от двух видов, не могли правильно соединяться в пары, и поэтому зрелые половые клетки оказывались содержащими различные числа хромосом и были неполноценными. Этот гибрид был поэтому стерилен. Но у него появилась ветка с удвоенным числом хромосом. Несомненно, что эта ветка возникла благодаря тому, что протоплазма одной клетки не разделилась после разделения хромосом, в результате чего получилась тетраплоидная клетка, имеющая по две хромосомы каждого сорта от обоих видов. Из этой клетки и возникла тетраплоидная ветка. На этой ветке образовались цветы, которые были фертильными, потому что хромосомы могли теперь соединяться в пары, ибо хромосомы одного вида соединялись с им подобными, а хромосомы другого вида тоже имели себе партнеров. Потомство оказалось тетраплоидным гибридом, и полученный новый вид плодовитым.
Если тетраплоид скрещен с диплоидом, то потомство будет триплоидным и, как правило, имеет не полноценные половые клетки. Потомство это стерильно благодаря тому, что все три сходные хромосомы не могут соединиться в пары, что при созревании половых клеток приводит к неправильностям в распределении хромосом. Случается, однако, что половая клетка может получить гаплоидный набор хромосом. Если соединяются две такие клетки, то диплоидное состояние опять восстанавливается. Другие возможные комбинации дают неуравновешенные типы потомков.
Как было описано выше, тетраплоидные клетки могут появляться у растения путем неразделения протоплазмы при расхождении хромосом. И действительно, в растительных тканях иногда встречаются тетраплоидные клетки. Такие клетки могут быть получены искусственно, путем воздействия на клетку во время процесса деления, агентом, препятствующим следующему клеточному делению. Дочерние хромосомы образуют тогда одно ядро с удвоенным числом хромосом, и это число сохраняется в течение всех последующих делений. Изредка могут также возникнуть и половые клетки с диплоидным числом хромосом вместо гаплоидного. Не может быть сомнений, что такие клетки получаются в результате подавления деления протоплазмы. {124} Такие тетраплоидные половые клетки крупнее нормальных.
Естественно, напрашивается вопрос, останется ли клетка крупнее после дальнейших делений, если к ней искусственно прибавить протоплазмы; и если это так, то останутся ли теми же размеры хромосом. В действительности было показано, что на некоторых стадиях дробления, после того как клетка разделилась на одну большую и другую меньшую, хромосомы в последней мельче, чем в первой. Являются ли гены затронутыми таким же образом, неизвестно. В настоящее время на этот вопрос не может быть дано ответа, но если бы общее количество протоплазмы могло быть искусственно увеличено, то это служило бы доказательством наличия в ней саморазмножающихся единиц, не зависимых от генов. Так как известно, что гены влияют на общее количество протоплазмы, как, например, это имеет место у тетраплоидов, то ясно, что между генами и протоплазмой существует некоторое взаимоотношение и, нужно думать, что предложенный выше опыт в случае отрицательного результата не пролил бы никакого света на этот вопрос.
Имеются некоторые данные об обратном положении вещей, когда гаплоидные особи или гаплоидные клетки появляются у диплоидной особи. В этом случае у гаплоидных растений отдельные клетки, повидимому, мельче, чем у диплоидов. У гаплоидных самцов животных, подобных коловраткам, тлям и пчелам, имеющиеся в настоящее время данные недоказательны. Гаплоидные участки случайно были найдены у диплоидных особей дрозофилы. Клетки в таких участках мельче. Является вероятным, что в других случаях общее количество протоплазмы коррелировано с числом хромосом и может показаться, что количество протоплазмы зависит от генов, но вопрос о том, имеются ли в протоплазме саморазмножающиеся единицы, которые наследуются независимо от единиц в хромосомах, или же увеличение, или уменьшение количества протоплазмы зависит от генных отношений, все же остается открытым.
| << | {125} | >> |
Относительная роль генов и протоплазмы в раннем и позднем развитии признаков индивидуума является одной из центральных проблем как генетики, так и эмбриологии.
Наиболее важные выводы в этой области вытекают из работы с реципрокными гибридами гермафродитных растений, у которых при скрещивании в одном направлении привносится протоплазма яйца одного вида, а при противоположном — другого. Оба гибрида содержат одинаковые гены, половина которых происходит от одного родителя, а половина — от другого. Так как во многих случаях взрослые гибриды от обоих скрещиваний идентичны, то из этого следует, что окружающая хромосомы протоплазма является индиферентной, по крайней мере, до момента достижения стадии взрослого организма. Таким образом, признаки, образуемые протоплазмой, определяются генами. Однако проблема является несколько более сложной потому, что в некоторых случаях в протоплазме яйца известны саморазмножающиеся элементы. Имеются, например, растения (Pelargonium), у которых случайно попадаются белые листья, объясняющиеся отсутствием в них хлорофильных зерен. Эти зерна нормально присутствуют в виде бесцветных пластид в яйце зеленого растения, но не в пыльцевой трубке, а если и присутствуют в пыльцевой трубке, то не переносятся пыльцевым ядром в яйцо. В яйцах, происходящих из белых частей белого растения, эти пластиды отсутствуют. Если пыльца с белой ветки оплодотворяет яйцо зеленого растения, то возникает зеленое растение. Наоборот, если пыльца с зеленого растения оплодотворяет яйцо с белой ветки, то возникающее растение оказывается белым. Пластиды, являющиеся предшественниками хлорофильных зерен, передаются в этих случаях только через протоплазму яйца. {126}
Прекрасной иллюстрацией частичной зависимости элементов протоплазмы от генетической конституции являются реципрокные гибриды между двумя видами «ночной красавицы» Oenothera muricata и О. suaveolans. Пластиды suaveolans не могут образовывать зэленый хлорофил в генетической среде гибрида и поэтому, когда материнским растением является suaveolans, проростки оказываются белыми и погибают. У реципрокных гибридов, где матерью является muricata, материнские пластиды могут развивать зеленый пигмент в гибридном генотипе, который, конечно, такой же самый, как и в первом скрещивании, и молодые растения оказываются зелеными.
К сказанному выше необходимо сделать небольшую оговорку. Изредка небольшое число пластид, повидимому, может все-таки проникать в яйцо вместе с пыльцевыми ядрами. Это обстоятельство усложняет результат лишь в том отношении, что, благодаря неправильностям в распределении этих пластид при первых делениях яйцевых клеток, может возникнуть небольшое число зеленых и белых мозаиков.
Конечно, в протоплазме могут быть и другие саморазмножающиеся тела, но до сих пор они неизвестны, а если даже в исключительных случаях они и существуют, то они не оказывают серьезного влияния на сделанное только что общее заключение, основанное на результатах возвратных скрещиваний.
Дополнительные данные дает экспериментальная эмбриология. Самые первые и простые из них основаны на скрещиваниях между различными видами морских ежей. Так, например, темп первого дробления яйца различен у разных видов. Если яйцо одного вида оплодотворяется спермой другого, то оно дробится со скоростью, свойственной тому виду, к которому оно принадлежит. Яйца одних видов содержат больше пигмента, чем яйца других. Молодые гибридные личинки содержат то же количество пигмента, которое было в материнском яйце, причем этот пигмент личинка получает непосредственно из неоплодотворенного яйца. Однако на более поздних стадиях при развитии нового пигмента можно увидеть влияние хромосом, содержащихся в сперматозоиде. Число мезенхимных клеток различно у разных видов. У Sphaerechinus, например, их 38, а у Echinus 57. Гибрид же, происходящий из яйца Sphaerechinus, имеет около 38 клеток мезенхимы. Одним из наиболее резких примеров относительного влияния {127} яйца и сперматозоида является скрещивание между Cidaris и Lytechinus. В следующей таблице дается длительность протекания различных стадий в часах.
Cidaris |
Часы |
Lytechinus |
Часы |
Бластула (плавающая) |
16—18 |
Бластула (плавающая) |
5,5 |
Гаструла (начало |
20—23 |
Мезенхима |
8 |
Мезенхима |
23—26 |
Гаструла (начало |
9 |
Хроматофоры |
44 |
Хроматофоры |
15—16 |
Скелет (начало |
72—73 |
Скелет (начало |
15—16 |
Плутеус |
120 |
Плутеус |
24 |
При оплодотворении яиц Cidaris спермой Lytechinus скорость дробления равна таковой у Cidaris; бластула тоже подобна бластуле Cidaris. У Cidaris мезенхимные клетки развиваются из внутреннего конца первичного кишечника, тогда как у Lytechinus они начинают мигрировать внутрь до начала гаструляции. Мезенхимные клетки гибрида возникают по краям и вокруг основания первичного кишечника, а не из внутреннего конца, как у Cidaris. Представляется поэтому, что на место возникновения мезенхимных клеток влияет сперматозоид Lytechinus.
Форма молодой треугольной личинки, получающейся при оплодотворении яиц Sphaerechirius сперматозоидами Strongylocentrotus, подобна таковой материнского вида, но на стадии плутеуса, которая следует непосредственно за стадией треугольной личинки, становится видимым влияние отцовских хромосом. Имеются многочисленные описания более взрослых стадий плутеусов гибридов между различными видами морских ежей, и практически у всех из них могут быть обнаружены характерные черты обоих родительских типов. Немногочисленные попытки произвести генетический анализ признаков (таких, как форма игольчатого скелета, форма личинки, характер пигмента и т. д.) не были очень удачными, так как хотя и оказалось возможным установить доминирование одного признака над другим, но число участвующих факторов не может быть определено без получения еще одного поколения от гибридов. Только таким путем, {128} как и в большинстве случаев генетического анализа, из числовых отношений между вновь появляющимися типами может быть определено число участвующих в скрещивании факторов.
Общие выводы из этих данных ясны. Ранние стадии развития определяются протоплазмой яйца; влияние же хромосом сперматозоида сказывается, только позже. Это означает, что протоплазма яйца уже подверглась влиянию генов, содержащихся в самом яйце, и требуется время для того, чтобы отцовские гены вызвали изменение в протоплазме клеток эмбриона.
Опыты по гибридизации усложняются двумя другими моментами, именно влиянием среды и неправильностями в распределении хромосом, которые иногда имеют место на ранних стадиях дробления гибрида.
Действие температуры изучалось как на чистых, так и на гибридных плутеусах, причем некоторое влияние ее было обнаружено. У гибридов, повидимому, от температуры зависит большее или меньшее доминирование материнского или отцовского влияния. Щелочность или кислотность морской воды в некоторых случаях тоже, повидимому, влияет на результат.
В некоторых скрещиваниях было показано, что на первых или более поздних стадиях дробления яйца может иметь место элиминация единичных или даже нескольких хромосом. Так, при оплодотворении яиц Arbacia сперматозоидами Sphaerechinus могут быть элиминированы около 18 хромосом предположительно отцовского происхождения, что может объяснять преобладание материнских черт у гибридного плутеуса. В реципрокном скрещивании не происходит элиминации и тем не менее плутеус оказывается сходным с таковым материнской расы. Элиминация хромосом, нарушающая сбалансированный эффект генов, может иметь и дальнейшее влияние, вызывая у эмбриона патологические признаки.
Для гибридизации многократно использовались также костистые рыбы, отдаленные виды которых могут широко скрещиваться друг с другом, но только в некоторых скрещиваниях эмбрион развивается до взрослой стадии. Скорость дробления при этом равна скорости дробления у материнского вида за исключением случаев, когда она замедляется вследствие ненормального разделения хромосом. Но при скрещивании близко родственных видов было {129} установлено, что темп дробления может быть ускорен отцовскими генами, если отец принадлежал к типу с более быстрым дроблением. Развивающиеся пигментные клетки происходят не из пигмента, содержащегося в яйце, а пигмент развивается в них независимо и может быть подвержен влиянию отцовских генов. Элиминация хромосом описывалась в различных скрещиваниях. Многие эмбрионы погибают на ранних стадиях, что, вероятно, в одних случаях объясняется элиминацией и неправильными делениями хромосом, в других же это более вероятно объяснить несовместимостью эффектов генов обоих видов.
 |
|
Рис. 76. Схема материнского наследования у шелковичного червя. |
У молодых эмбрионов некоторых рас шелковичного червя пигмент появляется в эмбриональной мембране из веществ, предшествующих ему в яйце. При скрещивании рас с различно окрашенной эмбриональной мембраной цвет ее у потомства оказывается материнским. Когда из этих яиц были получены взрослые гибридные бабочки, то было найдено, что у их эмбрионов развился доминантный цвет независимо от того, был ли он привнесен отцом или матерью (рис. 76). Что здесь мы действительно имеем случай генной наследственности, было показано путем выведения еще одного поколения, в котором эмбрионы (F.) расщепились в отношении 3:1 (или более точно 1:2:1). Этот результат показывает, что эмбриональный признак {130} определяется генами, которые следуют менделевскому закону расщепления для единственной пары факторов (рис. 76). Другими словами, материнская наследственность принципиально не отличается от обычной менделевской. В этом случае предшественники пигмента уже заложены в яйце, благодаря влиянию генов в течение ранних стадий формирования яйца, и не сразу изменяются под действием отцовских генов, даже если последние доминантны. Но в этом случае эффект доминантных генов проявляется в следующем (F2) поколении. В этом поколении имеются три генетически различных класса особей; доминантные и
 |
|
Рис. 77. Схема образования эндосперма у кукурузы. |
рецессивные гены расщепились в F1 и каждый, действуя на развитие протоплазмы своего собственного яйца, образует в F3 тот или иной тип эмбриона.
У этой бабочки имеются некоторые расы, дающие две генерации в год, тогда как другие расы имеют только одну. Было показано, что это различие тоже обнаруживает материнское наследование и вне всякого сомнения обусловлено какой-то особенностью яйца или яйцевой мембраны, которая в свою очередь обусловлена генами.
Подобные же явления найдены у индийской кукурузы при образовании эндосперма семян. Эндосперм является соматической тканью, образующейся в зародышевом мешке — увеличенной клетке — путем соединения двух материнских {131} ядер и одного отцовского. Цвет эндосперма определяется доминантными и рецессивными генами, содержащимися в этих ядрах. Например: если в качестве материнского растения была взята мучнистая кукуруза (ff), а в качестве отцовского кремнистая, то эндосперм получится мучнистым (ff F), откуда следует, что две дозы или два гена мучнистости доминируют над одним геном кремнистости (рис. 77). Наоборот, если кремнистая кукуруза была материнским растением, а мучнистая — отцовским, то эндосперм получается кремнистым (FFf), так как два гена кремнистости доминируют над одним мучнистости. Правильность этих формулировок показана выведением следующих поколений, в которых оба признака эмбриона (F и f) расщеплялись как одна пара генов. Надо отметить, что в этих двух случаях протоплазма эндосперма не изменялась геном пыльцевого зерна, потому что два рецессива подавляют собой доминантный ген.
Наконец, интересный случай материнской наследственности был найден у некоторых пресноводных моллюсков (Lymnaea). Раковины этих моллюсков обычно закручены в правую или декстральную спираль, но случайно встречаются формы, имеющие левые или синистральные витки спирали. Все потомство данного выводка, включая гибридов, имеет либо декстральные, либо синистральные витки. Было также показано, что некоторые матери с синистральными витками дают только синистральные семьи, тогда как другие такие же самки — только декстральные. Наоборот, некоторые декстральные матери производят только синистральные семьи, в то время как другие могут давать декстральное потомство. Эти факты были сначала с генетической точки зрения очень загадочными, но теперь мы имеем для них удовлетворительное объяснение. Предположим, как на это указывают факты, что имеется доминантный ген для декстральных витков и рецессивный для синистральных и что гены эти содержатся в данной паре хромосом. Самооплодотворяющаяся декстральная улитка гетерозиготная по этим генам (Ll) образует при созревании два сорта яиц L и l. Подобным же образом будет два сорта сперматозоидов L и l. Самооплодотворение приведет к образованию трех генетически различных типов потомков — LL, Ll и ll, но все эти особи будут иметь декстральные завитки, потому что тип дробления был определен уже в яйце доминантным фактором L еще до выделения {132} полярных телец. Из этих трех типов первые два (LL и Ll) будут давать только декстральное потомство, а третий тип (ll), также имеющий декстральные витки, даст только сини-
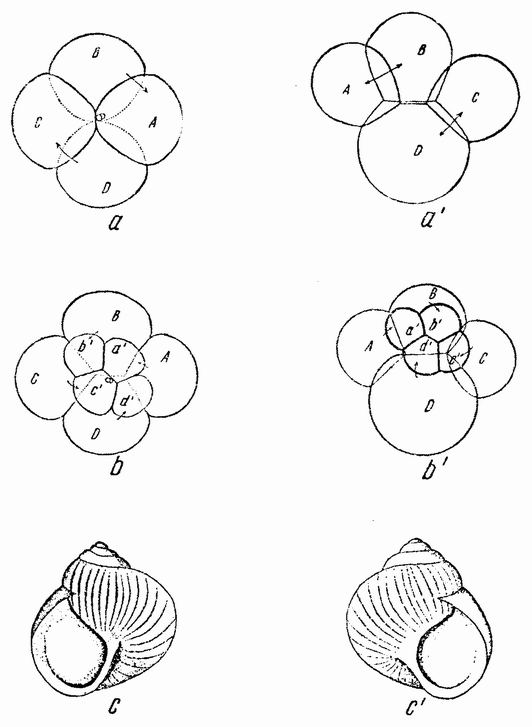 |
|
Рис. 78. а, b, с — левый тип дробления и влево закрученная спиральная раковина моллюска; а1, b1, с1 — правый тип дробления и вправо закрученная спиральная раковина моллюска (по Конклину). |
стральных потомков. Так как эти улитки могут оплодотворяться также и перекрестно, то при скрещивании декстральной улитки с декстральной или синистральной с {133} синистральной декстральная самка генетического строения ll (возникшая указанным выше путем) будет иметь возможность скреститься с декстральной же, но строения LL. Все потомство такой соматически декстральной самки будет синистральным, так как до начала дробления яйцо подвергалось влиянию двух рецессивных генов (ll). Эти синистральные улитки (Ll) в свою очередь будут давать только декстральных потомков, потому что доминантный фактор L в яйце определяет тип дробления последнего. Очевидно поэтому, что декстральные особи определенного
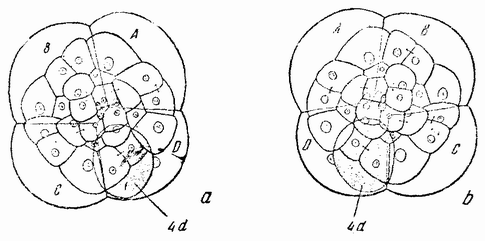 |
|
Рис. 79. а — более поздняя стадия дробления яйца моллюска, в которой с правой стороны обособилась 4d клетка; b — то же самое с 4d клеткой с левой стороны. |
происхождения будут давать только синистральные семьи, а некоторые синистральные — только декстральное потомство. Наследование здесь идет согласно Менделю, но проявление признака запаздывает на одно поколение. Результат этот своеобразен потому, что симметрия взрослого организма определяется не его собственной генетической конституцией, а конституцией еще несозревшего яйца, из которого он возник. Здесь нет противоречий с обычным менделевским наследованием признаков взрослого организма, если, а это кажется правильно, симметрия определяется строением яйца перед выделением полярных телец и будучи однажды детерминированной, не может впоследствии измениться в обратном направлении независимо от того, какое генетическое строение будет иметь зигота. Что яйцо построено именно так, видно из первых стадий дробления. На четырех- и восьмиклеточной стадиях спираль дробления имеет одно направление у влево закрученных (рис. 78 a, b, c) и противоположное у вправо закрученных {134} рас (рис. 78 а1, b1, с1). Когда появляется мезодермальная клетка (4d), то она оказывается с одной стороны от проспективной средней линии у одного типа и с противоположной — у другого (рис. 79 а, b).
Все генетические данные приводят к выводу, что признаки, развивающиеся в протоплазме, в конце концов оказываются зависящими от генов в хромосомах. Более того, они показывают, что протоплазма яйца, образовавшаяся под влиянием одного набора генов, может со временем быть измененной в протоплазму, характерную для другого набора генов, привнесенного ядром сперматозоида. В случае реципрокных гибридов видно, что два набора хромосом, происходящих от двух различных рас, вызывают один и тот же конечный результат (если имеется достаточно времени для их действия) независимо от того, какая протоплазма присутствовала сначала.
С другой стороны, имеется несколько случаев, в которых реципрокные гибриды не идентичны. Кроме случаев элиминации хромосом в одних скрещиваниях и отсутствия ее в других, можно иногда ожидать различий другого рода. Так, например, случай с моллюсками показывает, что очень ранние стадии дробления по своим признакам оказываются материнскими.
Если бы эти начальные стадии влияли на все или некоторые из более поздних, то можно было бы предположить, что как своего рода последействие начальных стадий получится различный конечный результат. Если это так, то реципрокные гибриды могли бы быть различными, что и имеет место в действительности в некоторых скрещиваниях с вправо и влево закрученными улитками. Точно так же у млекопитающих, у которых плод получает питание от матери, можно было бы ожидать, что реципрокные гибриды обнаружат при рождении различия, обусловленные различиями в среде, в которой они развивались.
В связи с этим остается обсудить довольно существенный вопрос. Можно ожидать,что только в тех случаях оказывается возможным получение реципрокных гибридов, достигающих взрослой стадии, в которых протоплазма обоих родителей в. основном подобна, поэтому рискованно переносить доказательства, вытекающие из этих фактов, на все остальные случаи. Ибо если имеются реальные различия в протоплазме различных видов, не зависящие от ядерных влияний в том смысле, что они не могут быть обращены {135} одно в другое влиянием генов, то нельзя ожидать развития организма до взрослого состояния или до последних эмбриональных стадий. Поэтому до тех пор, пока не получены более обширные данные, затронутая центральная проблема не может считаться разрешенной для всех случаев.
Вопрос об относительной роли генов и протоплазмы, иллюстрированный вышеприведенными примерами, соприкасается с другим вопросом, дискутировавшимся в прошлом, а именно, что протоплазма определяет основные признаки особи, а гены только более поверхностные из них. Необходимо лишь напомнить, что опыты с влево и вправо закрученными улитками показали, что гены могут в течение времени изменить одну форму протоплазматической реакции в другую. Если мы попытаемся определить, какой из этих признаков является основным, а какой поверхностным, то наверное реверзия симметрии будет с уверенностью отнесена к первой категории признаков. Наследование четырех групп крови у человека, составляющих тройную аллеломорфу, будет также отнесено к случаям, в которых основной признак определяется генным влиянием. Наоборот, если пигмент рассматривать как поверхностный признак, то случай с шелковичным червем показывает, что он тоже может определяться протоплазмой яйца, но в последующих поколениях может быть изменен влиянием генов.
В течение долгого времени было известно, что при некоторых условиях уже диференцировавшиеся клетки, выполнявшие определенные функции во взрослом индивидууме, могут полностью или частично передиференцироваться и затем играть другую роль в построении новой особи. Так, например, ткани некоторых асцидий в определенное время года могут образовывать шарообразные вздутия, состоящие из вещества, в котором исчезли все признаки взрослой диференцировки и из которого впоследствии может развиться новая особь. Здесь нет ясной непрерывности между типом клеток старых и новых органов. Ткань губок и гидроидов может быть протерта через мельничный газ и если затем она будет помещена на дно сосуда с морской ведой, то в результате соединения клеток, разъединенных протиранием, может образоваться {136} несколько новых особей. В какой степени любая клетка может быть частью нового организма или в какой мере ее первое состояние может влиять на ее последующую судьбу, не вполне ясно, но тем не менее в некоторых клетках должна произойти большая передиференцировка.
Некоторые опыты с прививками частично дают обратную картину. Если кусок кожи со спины только что вылупившегося цыпленка, принадлежащего к породе, отличающейся особенностями оперения, трансплантиру-
 |
|
Рис. 80. Недавно привитые цыплята (по Депфорду). |
ется на то же место цыпленку другой породы (рис. 80), кожа которого в этом месте предварительно удалена, то новые перья привитого участка оказываются подобными перьям той породы, от которой происходит привитой кусочек (рис. 81). Во время операции у цыпленка имеется только пух, но фолликулы перьев уже заложены и, как установлено выше, на новом хозяине они дают начало тому же самому типу перьев, который бы они выработали, если бы остались на своем месте. Более того, если они будут выдернуты или если они вылиняют, то развивающиеся новые перья происходят из новых фолликул, образованных из старых, и сохраняют свой тип. Здесь генетическое строение отдельных клеток определяет признаки оперения привитого участка кожи, даже когда он оказывается лежащим во внешней среде хозяина, имеющего {137} совершенно отличные перья. Передиференцировки в этих условиях не происходит.
С другой стороны, было показано, что некоторые черты оперения подвержены влиянию внешних условий или специфичному воздействию внутренней секреции хозяина.
 |
|
Рис. 81. Взрослые куры с прививками на крестце, а — красная курица с прививкой от петуха попосатого плимутрока; b — петух род айленд с прививкой от петуха полосатого плимутрока; с — петух рыжий лехгорн с прививкой от джерсейской белой гигантской курицы; d — курица, белый лехгорн с прививкой от курицы красный род айленд (по Денфорду). |
Так, например, если кожу с крестца молодого самца плимутрока привить на красную самку, то перья оказываются подобными соответствующим перьям самки плимутрока, а не самца (рис. 81 а). Другими словами, внутренняя секреция самки определяет некоторые признаки нового оперения. Крестцовые перья у петухов длинные, узкие и {138} заостренные, у самок же они в этом месте короткие и закругленные.
Обратная прививка обнаруживает сходный эффект. Если, например, кусок кожи с крестца черной курицы привит на спину красно-желтого петуха лехгорна, то новое черное оперение будет подобным длинным заостренным перьям петуха (рис. 81 с). Здесь этот результат объясняется отсутствием внутренней секреции, имевшейся у самки, что видно из того, что при удалении у самки яичника, ее новые крестцовые перья становятся подобными перьям самца.
Эти результаты согласуются с точкой зрения, что в случаях, когда протоплазма однажды уже была диференцирована, конечный продукт этим определен даже тогда, когда новые клетки образуются из исходных клеток, так сказать, уже направленных по определенному пути на ранней эмбриональной стадии. Результаты, касающиеся признаков окраски оперения, показывают, казалось бы, что гены не были изменены. С другой стороны, поведение в этих опытах признаков формы перьев показывает, что внутренняя среда, т. е. гормоны, вырабатывающиеся в других частях тела, тоже действует на некоторые конечные стадии диференцировки. Гормоны могут действовать сначала на протоплазму фоликулярных клеток перьев и далее посредством вызванного таким образом эффекта на продукт гена, или же они могут действовать непосредственно на гены. В настоящее время нет путей для определения, какое из этих воздействий имеет место в действительности. Имеющиеся данные не являются также решающими для вопроса, действуют ли все гены одинаково или одни из них активнее других.
Сравнение эмбрионального развития цветных пятен у пегих животных с наследованием этого же самого цвета, свойственного другим расам с одноцветной окраской шерсти, может быть обманчивым. Существуют морские свинки серые (агути), черные, красные и белые, причем наследование этих окрасок объясняется различиями в единственном гене. Имеются также пегие морские свинки. Некоторые из них могут иметь одновременно серые и белые участки шерстного покрова, другие — красные и белые, третьи — черные и белые. На первый взгляд может показаться парадоксальным, что морская свинка, у которой могут развиваться черные участки, будет иметь белые пятна в случае, если клетки обоих участков будут содержать все гены. В {139} тех случаях, где пятна точно локализованы, т. е. распространяются на определенные области тела, их локализация может не казаться отличной от какой-либо другой соматической локализации; но когда, как это имеет место у некоторых пегих животных, черные и белые пятна распределяются случайно, отмеченный выше парадокс более очевиден.
Генетические данные показали, что у мышей, крыс и морских свинок имеются определенные гены пегости. Когда присутствуют два таких рецессивных фактора, животное оказывается пегим, т. е. оно имеет участки белого и какого-либо другого цвета; например, серого. Согласно нашему предположению, гены серого цвета присутствуют во всех клетках; почему же тогда серый цвет проявляется не везде? Этот вопрос требует рассмотрения.
Для образования пигмента должно присутствовать по крайней мере два вещества; если одно из них отсутствует в клетке или не вырабатывается в ней, то окраска не развивается. В присутствии генов пегости ген серого цвета в белых участках теряет способность вырабатывать необходимый для этого цвета энзим или же протоплазма клеток некоторых участков тела не соответствует продукту таких генов. И та, другая альтернатива приводит нас к, повидимому, неразрешимой дилемме, ибо при первом предположении нельзя объяснить что подавляет ген, а при втором, повидимому, необъяснимо, почему эти протоплазматические различия находятся не на всем теле.
Эти трудности объяснения не встречаются по отношению к чисто белым но тем не менее содержащим гены серой, красной или черной окраски животных. Здесь предполагается, что в присутствии нормального аллеломорфа рецессивного фактора белого цвета вырабатывается одна из субстанций, необходимых для развития окраски. Рецессивный фактор этого свойства лишен, и развития окраски поэтому не происходит.
Здесь надо подчеркнуть, что фактор пятнистости вызывает свой эффект не тем же путем, что и фактор одноцветной белой окраски, поэтому незаконно постулировать, что взаимоотношения энзимов в обоих случаях одинаковы; в действительности они должны быть различными. Следовательно, вопрос о том, каким образом фактор пятнистости вызывает свой локализованный эффект, остается еще открытым. {140}
Существуют некоторые опыты по прививкам участков кожи у морских свинок, показывающие, что если эффект однажды был вызван, то результат не может быть изменен в обратном направлении путем перемены местоположения белого и черного участков. Когда участок кожи с черной шерстью переносится на белый участок той же самой свинки, шерсть остается черной; наоборот, белый участок на черном тоже остается белым. Тот же результат получается с черным и серым, серым и красным и т.д. Очевидно, здесь уже не локализация пятна, все равно белого или черного, определяет его цвет, ибо, будучи однажды определенным, он оказывается зафиксированным навсегда.
Это еще раз ставит вопрос, действительно ли диференциация черных и белых участков была определена их локализацией в эмбрионе или какими-либо диференциаторами, вроде, например, саморазмножающихся протоплазматических элементов, неправильно распределившихся во время раннего развития. Это приводит к положению, вновь возвращающему нас к проблеме локализации. Например, у всех серых морских свинок шерсть на спине совершенно отлична как по цвету, так и подлине от шерсти на брюхе. Если участок кожи с брюха пересажен на спину, он сохраняет все признаки брюшной шерсти. Если, как это обычно предполагают, различия между спиной и брюхом являются проблемой эмбриональной локализации, то почему мы должны воздерживаться от пользования этой же аргументацией для пегих животных, хотя бы даже, вследствие неправильности в распределении пятен, проблема казалась бы несколько более трудной?
| << | {141} | >> |
У всех крупных групп животных, молодь которых развивается как в пресной, так, в особенности, в морской воде, имеются типичные для каждой группы свободно плавающие личинки, способствующие широкому распространению видов. Для завершения своего развития они должны добывать себе пищу, так как они происходят из мелких яиц, не содержащих достаточно желтка, чтобы довести личинку до взрослой формы. Типичным личинкам каждой группы присвоено специальное название. Хотя личиночная стадия каждого вида обнаруживает свои характерные признаки, тем не менее внутри группы личинки имеют сходную основную структуру. В этой главе будут описаны некоторые из наилучше изученных типов личинок и обсуждена интерпретация их.
У иглокожих морские ежи проходят стадию плутеуса; другие группы — морские звезды, морские лилии и голотурии — тоже имеют довольно сходные и характерные для них личинки. Плутеус (рис. 82) представляет собой небольшую прозрачную личинку с известковым скелетом, снабженную на своей поверхности довольно сложным рядом реснитчатых клеток, при помощи которых плутеус активно плавает в море. На одной стороне личинки находится широкий рот, ведущий в трубкообразный пищевод, который открывается в большой желудок. Последний переходит в короткий кишечник, открывающийся с дорзальной стороны. Между кишечником и внешней стенкой имеется пространство, заполненное жидкостью и содержащее свободные мезенхимные клетки, некоторые из которых образуют известковый скелет. Кроме того, на дорзальной стороне имеется небольшой мешок, открывающийся наружу с помощью небольшой трубки. Стенки этого мешка образуют впоследствии внутренний слой полости тела и {142} гастро-васкулярную систему. Когда личинка вполне развилась, в ней происходит внезапное изменение. Вокруг рта появляется масса клеток, происходящих из личиночных органов, которые превращаются в весьма отличную форму молодого морского ежа или звезды. Старый покров медленно всасывается внутрь растущего ежа, а личиночный скелет исчезает. Такое резкое и внезапное изменение формы, подобное только что описанному, называется метаморфозом. Эмбрион опускается на дно, причем его движения
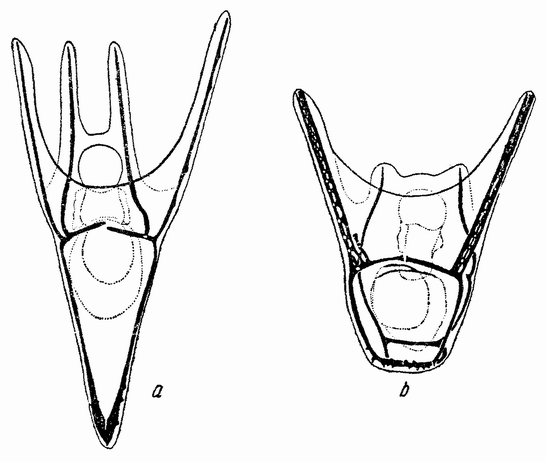 |
|
Рис. 82. а — плутеус Strongylocentrotus; b — плутеус Sphaerechinus (по Гербсту). |
совершаются теперь с помощью системы трубчатых ножек, располагающихся вдоль пяти лучей вновь образованного тела. Личинка имела двустороннюю симметрию, взрослое же животное радиально симметрично.
Морские кольчатые черви и моллюски проходят свободноплавающую стадию, известную под именем трохофоры (рис. 83 а). Трохофора имеет два ряда ресничек, а на своем переднем конце несет группу ресничек, в основании которых находится очень примитивный нервный центр. Пищеварительный тракт состоит из рта, пищевода, желудка, {143} кишечника и заднепроходного отверстия. Между наружной стенкой пищеварительного тракта и внутренней стенкой тела имеется большое пространство с немногочисленными мускульными клетками, тянущимися вокруг и вдоль стенок. У кольчатых червей на заднем конце имеется небольшая группа мезодермальных клеток, из которых развиваются будущие сегменты и мускулатура. Превращение трохофоры в червя происходит постепенно. Задний конец трохофоры вытягивается (рис. 83 b); из мезо-
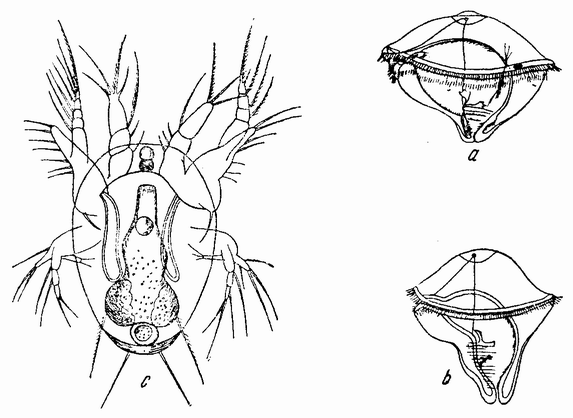 |
|
Рис. 83. а — трохофорная личинка кольчатых червей; b — более поздняя стадия ее, обнаруживающая удлинение заднего конца и развитие сегментов: с — личинка ракообразных — науплиус (по Гатчеку из Коршельта и Гейдера). |
дермы одна за другой образуются парные группы клеток. Каждая пара содержит полость, внутренние стенки которой образуют мускулатуру пищеварительного тракта, а наружные — мускулатуру стенки тела. Между располагающимися друг за другом сегментами образуется септа; головной конец редуцируется; мозг увеличивается, и эктодермальное утолщение вдоль вентральной стороны становится нервной струной.
Превращение трохофоры моллюсков происходит несколько отличным путем. В качестве выделения кожного {144} покрова на дорзальной стороне развивается раковина, которая впоследствии облекает животное, когда оно сокращается. Утолщение вентральной стенки образует мускулистую ногу. Взрослое животное не разделено на
 |
|
Рис. 84. Hydatina senta. A — взрослая самка; В — молодая самка вскоре после вылупления; С — взрослый самец; D — партеногенетическое яйцо; Е — яйцо, дающее самца; F — покоящееся яйцо (по Уитнею). |
членики, как аннелиды, а состоит из одного единственного переднего сегмента или во всяком случае имеет не более одного добавочного. Взрослые моллюск и аннелида эволюционировали совершенно различными путями.
Наличие у обеих групп общего типа личинки интерпретировалось как доказательство того, что эта личинка {145} является предком, из которого развились обе группы. Если принять в расчет другие соображения, то кажется более вероятным, что присутствие у обеих групп трохофоры скорее означает, что они произошли от общего предка, который проходил через сохранившуюся до сих пор стадию трохофоры.
Идентичность их типов дробления подтверждает этот взгляд, так как стадии дробления конечно не были взрослым предком.
Существует группа пресноводных и морских животных, так называемые коловратки (рис. 84 а), которые во многих отношениях напоминают трохофорную личинку (рис. 83). Развитие коловраток никогда не идет дальше этой стадии, но у них образуется пара яичников или семенников. Были высказаны предположения, что коловратки представляют собой задержавшуюся трохофорную стадию какой-то древней аннелиды (или моллюска), которая больше не достигает прежнего взрослого состояния.
Большинство низших ракообразных имеет маленькую свободноплавающую личинку с шестью конечностями, (рис. 83 с), называемую науплиусом, который одно время тоже считался предком, из которого развились все ракообразные. Имеется, однако, больше оснований рассматривать науплиус как сохранившуюся до наших дней личинку предков ракообразных.
Большинство низших насекомых проходит через последовательный ряд стадий, каждая из которых более совершенна, чем предыдущая. Каждая стадия заканчивается линькой, при которой сбрасывается наружная кутикула, мешающая дальнейшему росту. Тем временем под старой кутикулой развивается новый мягкий покров. Заметное увеличение величины личинки происходит немедленно, прежде чем новая кутикула затвердевает. У наиболее высокоорганизованных групп насекомых — жуков, бабочек, мух, пчел и муравьев — имеется характерная личиночная стадия, сильно отличающаяся от взрослой формы. Организм на этой стадии называется личинкой или гусеницей. Переход от личинки к взрослой стадии у этих групп происходит внезапно. Совершается типичный метаморфоз. Между личинкой и взрослой формой находится неподвижная стадия, так называемая куколка, у которой внутри пуппария из дисков недиференцированных клеток, так называемых имагинальных дисков, формируются органы взрослого {146} животного. Большинство личиночных органов при этом полностью теряется и рассасывается.
Не пытаясь описывать метаморфоз насекомых, будет достаточным указать, что типы личинок этих высших групп не могут считаться полностью соответствующими личинкам предков, хотя некоторые признаки последних и могли быть сохранены. Личинки являются сами по себе высоко приспособленными формами, которые, может быть, настолько отошли от исходного типа, насколько сами взрослые формы отошли от других членов прародительской группы. Например, гусеница бабочки грызет свою пищу с помощью роговых жвал, тогда как сама бабочка имеет трубку для всасывания нектара из растений. У гусеницы развивается новый комплект конечностей; сложные глаза у нее отсутствуют. Личинка и взрослый организм стали настолько специализированными, что постепенный переход одного в другое невозможен. Поэтому, когда гусеница достигает своей конечной стадии развития, она перестает двигаться, вьет вокруг себя кокон и из ее имагинальных дисков недиференцированных клеток образуются органы бабочки. Сказать точно, каким образом это приспособление произошло исторически, — трудно, но у некоторых других групп имеются переходные стадии, которые делают эту трудность преодолимой.
У позвоночных нет вполне типичной для всех них личиночной стадии, но у амфибий таким типом личинки является головастик лягушки. Он тоже, подобно гусенице, представляет сильно специализированную личинку и в конце личиночного периода претерпевает внезапный метаморфоз.
Головастик лягушки многими своими особенностями напоминает соответствующую стадию более простых групп амфибий — тритонов и саламандр — и так как личинки последних по всем своим признакам, за исключением величины, очень похожи на взрослые стадии, то может показаться, что головастик представляет собой взрослую саламандру, повторяющуюся в развитии лягушки. Это и будет приложением так называемого биогенетического закона Геккеля. Однако имеется много оснований для того, чтобы счесть последнюю интерпретацию ошибочной. Головастик не является рекапитулировавшей стадией саламандры, а стадией развития общей и для лягушки, и для саламандры.
Когда головастик вырастает до своих полных размеров, {147} что у различных видов может продолжаться от нескольких недель до одного года или больше, он прекращает питаться и в течение нескольких дней претерпевает ретрогрессивные изменения в одних из своих органов и прогрессивные в других. Хвост головастика укорачивается и его внутренние ткани рассасываются. Они пожираются фагоцитами или белыми кровяными клетками, которые несут продукты их распада назад в организм. Большая часть жаберного участка рассасывается. Пищеварительный тракт головастика, представляющий собой длинную свернутую трубку, во время метаморфоза укорачивается и стенки его утолщаются. Роговые зубы во рту исчезают и рот расширяется. Жаберные щели закрываются, а сами жабры рассасываются. Кровеносные сосуды, проходящие через жаберные дуги, частично рассасываются, частично же превращаются в некоторые сосуды взрослого животного. Легкие, которые развиваются к тому времени в виде пары мешковидных выростов пищевода, начинают функционировать, как только в них набирается через нос воздух. Происходят дальнейшие видимые прогрессивные изменения. Передние и задние конечности, присутствующие у головастика в виде небольших рудиментарных органов, внезапно начинают увеличиваться. До начала метаморфоза передние конечности очень малы, во время же него они быстро растут и пробиваются сквозь закрывающую их до этого времени складку. Задние конечности, которые длиннее передних, тоже быстро растут. Интересно отметить, что оба обратных процесса происходят одновременно: одни органы дегенерируют, тогда как другие растут.
Метаморфоз тесно связан с изменениями в щитовидной железе, являющейся органом внутренней секреции, вырабатывающим гормон (Thyroxin), т. е. вещество, абсорбирующееся из железы кровью и вызывающее некоторые из вышеописанных изменений в теле. При содержании головастика в воде с примесью йодистых соединений или путем инъекции сходных веществ переход к лягушке может совершиться задолго до обычного срока.
Одна из хвостатых амфибий, а именно мексиканский аксолотль, достигает половой зрелости, когда у нее еще присутствуют жаберные щели и жабры. Аксолотль разводится обычно в этом состоянии. Его взрослая форма — сухопутная саламандра — развивается только при исключительных условиях. Созревание и функционирование {148} половых клеток — яиц и спермиев — в то время, когда особь находится еще на стадии личинки, называется неотенией. Можно подозревать, что это явление имеется и у других представителей хвостатых амфибий, а также и у некоторых других групп животных. Например, асцидии проходят через свободноплавающую личиночную стадию (рис. 85 а), напоминающую головастика лягушки. Существует также
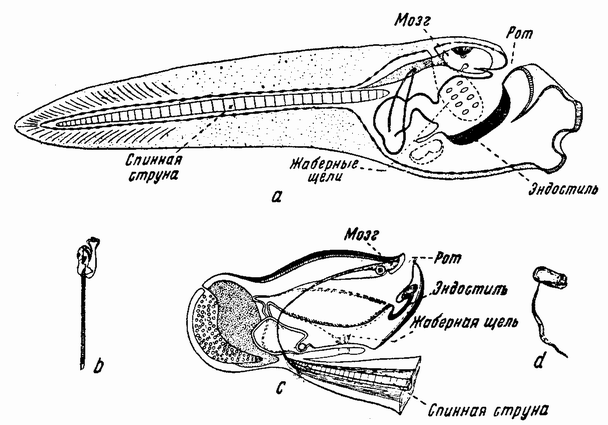 |
|
Рис. 85. а, b — стадия головастика у асцидии; с, d — свободноплавающая аппендикулярия. |
группа свободноплавающих пелагических асцидии, Append icularia (рис. 85, с), имеющая основные черты стадии головастика других асцидии. Эти аппендикулярии размножаются, но никогда не превращаются в форму, характерную для других членов этой группы. Они, вероятно, являются неотеническими формами.
В противоположность животным, обладающим многочисленными мелкими яйцами, дающими начало свободноживущим личинкам, существуют другие животные, образующие небольшое число крупных яиц, содержащих достаточно пищевых веществ для поддержания жизни зародыша в течение его последующего развития до той стадии, {149} которая всеми своими признаками, кроме размеров, напоминает взрослый организм. В этом случае можно сказать, что развитие является прямым. К зародышам с прямым развитием, которые до относительно поздней стадии своего развития находятся внутри материнского организма, применяется термин плод. Плод прикреплен к матке особым органом зародыша — плацентой, которая так соединяет кровеносную систему зародыша с кровеносной системой материнского организма, что зародыш абсорбирует из нее питательные вещества.
В большинстве крупных групп имеются виды с прямым развитием и эти виды обычно имеют очень крупные яйца.
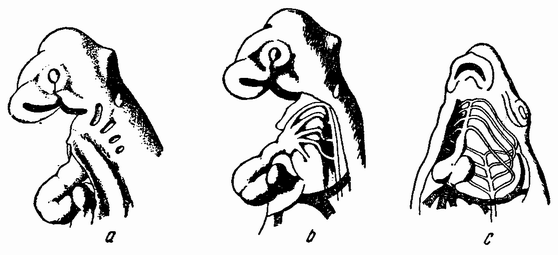 |
|
Рис. 86. а — голова цыпленка, обнаруживающая четыре жаберных щели (вид сбоку); b — дуги аорты; с — сердце и дуги аорты у рыбы (по Гессе). |
«Желток» куриного яйца в двадцать семь миллионов раз превышает объем яйца ланцетника. Наилучше изученные и наиболее интересные случаи известны у позвоночных. Огромные яйца имеют ящерицы и птицы, и молодь у них при вылуплении или при рождении имеет форму взрослого организма. Развитие млекопитающих весьма похоже на развитие птиц и ящериц, несмотря на тот факт, что оно берет начало из очень маленького яйца.
Сравнение развития этих типов друг с другом и с более низко организованными членами группы обнаруживает некоторые интересные отношения. Во-первых, наличие У ящериц, птиц и позвоночных (рис. 86 а, b) таких систем органов, как жаберные щели, которые напоминают эти же органы у свободноживущих личинок рыб (рис. 87) и амфибий; {150} во-вторых, присутствие эмбриональных новообразований, приспосабливающих зародыш к новым условиям, как, например, амнион и аллантоис (рис. 31). Кроме того, имеется много второстепенных признаков, относящихся к одной из этих двух категорий. Сравнение приводит нас назад к стадиям развития рыбы и амфибии. Рыбы, как хрящевые (акулы), так и костистые (Teleosti), дышат с помощью жаберной системы (рис. 86 и 87), состоящей из расположенных по бокам шеи жаберных щелей, которые внутри открываются в пищевод, а снаружи в воду. Между жаберными щелями находятся жаберные дуги, по которым кровь проходит от вентрального сердца к дорзальной аорте. На дугах име-
 |
|
Рис. 87. Голова молодой хрящевой рыбы сбоку; видны жаберные щели (по Седжвику). |
ется множество нитевидных жабер, содержащих капиллярные кровеносные сосуды. В жабрах кровь окисляется. Акт дыхания состоит в набирании в рот воды и в пропускании ее сквозь жаберные щели над жабрами. Эта система появляется в развитии рыб очень рано, независимо от того, происходят ли они из крупных яиц, как у акул, или из мелких, как у костистых рыб. Система эта, однако, не функционирует до вылупления молодого животного из яйца.
У эмбрионов ящериц, птиц и млекопитающих (рис. 88) на соответствующей стадии тоже появляются жаберные щели. Но они никогда не функционируют как органы дыхания, а скоро закрываются. На других жабры не образуются, но кровеносные сосуды, несущие кровь через дуги, функционируют с самого начала, неся кровь от сердца к спинной аорте. На более поздних стадиях некоторые из этих кровеносных {151} сосудов исчезают, другие же сохраняются, становясь частями кровеносной системы взрослого организма. Важными моментами являются при этом, во-первых, то, что у зародышей более высоко организованных групп дыхательный аппарат появляется на той же стадии, что и у более низко организованных групп; во-вторых, что у зародыша он выполняет функцию переносчика крови от сердца к спинной стороне; и в-третьих, что аортные дуги представляют собой структуру, из которой возникают более специализированные системы взрослого организма.
 |
|
Рис. 88. Человеческий эмбрион; видны сердце, дуги аорты и жаберные щели (по Гису из Маршаля). |
Амфибии представляют собой интересное промежуточное состояние. Они ближе, чем рыбы, к более высоко организованным типам, хотя они могут и не быть в числе их непосредственных потомков. Более того, некоторые амфибии являются одними из самых первых позвоночных животных, которые покинули воду, для того чтобы жить на суше, и которые обнаруживают переход от жабер к легким. Лягушки и многие хвостатые амфибии, живущие в настоящее время на суше, уходят для откладки яиц в воду, где протекает развитие их молодых стадий. Молодь сначала дышит при помощи эмбриональных наружных жабер, которые замещаются внутренними жабрами, подобными таковым у рыб. Жаберная система рассасывается, когда выросшие личинки покидают воду для сухопутной жизни, причем это рассасывание и трансформация в основном происходят так же, как и у птиц и млекопитающих. {152} Таким образом, ранние стадии молодых лягушек протекают так же, как и у воднодышащих рыб, а их взрослые стадии подобны таковым рептилий, птиц и млекопитающих.
Сходство молодых стадий высших групп позвоночных было подмечено и истолковано еще более ста лет назад, т. е. уже в то время, когда теория органической эволюции еще не была распространена. Этому сходству тогда было дано только формальное объяснение. Наиболее известное положение, именуемое в настоящее время законом Бэра, было установлено в 1828 году. Положение Бэра часто смешивается с биогенетическим законом Геккеля, который, однако, совершенно отличен и предполагает принятие эволюционной теории. Четыре закона Бэра таковы:
1. Наиболее общие признаки крупных групп появляются у эмбриона раньше, чем специализированные признаки.
2. За наиболее общими структурами возникают менее общие и так до тех пор, пока, наконец, не возникнут наиболее специализированные образования.
3. Эмбрион каждого данного взрослого животного не повторяет при своем развитии другие взрослые формы, а, наоброт, дивергирует с ними.
4. В основном эмбрион высшего животного никогда не бывает похож на взрослую стадию другой животной формы, а походит только на ее зародыш.
Так называемый биогенетический закон более определенен. Он гласит, что эмбрионы представителей высших групп проходят через взрослые стадии более низко организованных представителей этой же группы, которые как бы конденсированы в их развитии. Это положение так картинно изображает некоторые особенности развития, что оно стало одной из наиболее популярных теорий биологии. Его большая простота скрывает его запутанность и создает впечатление «объяснения», в то время как оно скорее служит для сокрытия исторического родства. Теперь, когда мы лучше понимаем, каким образом появляются новые наследственные изменения, мы находим, что нет постоянной связи между исходным признаком и появлением нового, как это подразумевается в биогенетическом «законе». Во многих случаях, может быть, даже в большинстве, новый конечный признак просто замещает исходный. Эмбрион развивает новый признак не после того, как он проходит через последнюю стадию исходного признака, хотя иногда это и может случиться, а новый признак занимает место {153} исходного. Так, например, мутант дрозофилы, имеющий рудиментарные крылья, при своем развитии не проходит через стадию с большими крыловыми пластинками или крупными дисками, а крылья непосредственно развиваются у него из дисков пропорционально уменьшенной величины. То же имеет место почти для всех известных мутантных форм, причем это верно не только для тех случаев, в которых новый орган оказывается уменьшенным, как в случае зачаточных крыльев, но и когда новый признак является добавлением чего-либо лишнего к тому, что было до этого.
Правда, имеются некоторые случаи, в которых новый признак налагается на старый, причем последний может до некоторой степени сохраняться. Например, некоторые жуки имеют черные пятна на желтом фоне. Известны черные мутанты, развитие которых проходит стадию, на которой присутствуют пятна на светлом фоне, но последний скоро темнеет, закрывая собой пятна. В этом случае можно сказать, что новый признак добавляется к старому в конце ряда, здесь, может быть, будет правильным сделать вывод, что взрослая стадия предка сохранилась в развитии последующей формы. Однако такие случаи являются исключениями. Правило же, как было установлено выше, состоит в том, что первоначальная конечная стадия выпадает из развития и замещается новым признаком.
Хотя Геккель и считал, что эмбриональные стадии по большей части подобны взрослым стадиям предков, но он был знаком также и с эмбриональными приспособлениями, которые за взрослые стадии никак не могут быть сочтены; таковы, например, аллантоис, амнион и желточный мешок птиц и млекопитающих. Он называл их фальсификацией филогенетически предшествовавших стадий. Они являются эмбриональными приспособлениями. Вскоре были найдены основания для скептического отношения к толкованию многих личиночных и эмбриональных стадий как предков ныне живущих форм. Таковы, например, науплиус ракообразных и трохофора кольчатых червей, которые рассматриваются теперь только как специализированные личиночные формы, и широкое распространение их в группе не есть достаточное доказательство того, что эти личинки соответствуют взрослым стадиям предков. Другими словами, если теория рекапитуляции является «законом», то он имеет так много исключений, что становится бесполезным и часто ошибочным. Более того, как было {154} установлено выше, он предполагает такой способ «прибавления» новых признаков, который не соответствует многочисленным фактам, которыми мы располагаем.
Теория гена с ее допущениями полностью согласуется со всем тем, что известно относительно развития вновь появляющихся признаков. Мутационное изменение в гене может быть таким, что его эффект становится видным только по достижении последней стадии развития, причем это изменение может эту стадию полностью изменить. Опять-таки может быть такого рода генное изменение, которое, влияя на личиночную стадию, изменяет один или большее число из ее признаков. Такое изменение может либо затрагивать взрослые стадии, либо нет. A priori должно казаться, что чем раньше совершается изменение в принятом типе развития, тем более вероятно, что оно будет затрагивать также и поздние стадии. Вся цепь явлений развития так сложна, что, вероятно, много труднее получить изменение в ранней стадии, не вызывая нарушения последних стадий, чем наоборот. Это обстоятельство могло создать благоприятные условия для сохранения ранних эмбриональных и личиночных стадий крупных групп.
Ни все законы Бэра, ни биогенетический закон Геккеля не находятся в соответствии с известными в настоящее время фактами изменчивости и развития. Возможно дать объяснение, много более простое и более согласующееся с этими фактами. Жаберные щели млекопитающих должны сравниваться с таковыми не у взрослых рыб, а с жаберными щелями молодых организмов на той же стадии. По мере эволюирования высших позвоночных из низших они сохранили много признаков начальных стадий своих предков. При желании можно было бы, став на телеологическую точку зрения, сказать, что либо этот путь является единственно «известным» для них методом достижения взрослой стадии, либо же, иначе, что они нашли, что наиболее простым способом для развития более сложных структур является пристройка их на тех, которыми организм пользовался уже миллионы лет. Такая точка зрения может быть названа принципом эмбрионального консерватизма. Выражаясь менее антропоморфично, этот принцип, которому не обязательно давать громкое название закона, а который может именоваться лишь принципом эмбрионального консерватизма, заключается в том, что высшие и низшие члены группы проходят одни и те же стадии развития.
| << | {155} | >> |
История партеногенеза одна из наиболее интересных глав биологии. Еще Аристотель подозревал, что некоторые касты медоносной пчелы размножаются без спаривания, но изображаемая им картина этого явления была большей частью неверной. До конца семнадцатого или до начала восемнадцатого века почти не было наблюдений, показывающих, что яйца некоторых бабочек могут развиваться и в тех случаях, когда самка не была спарена; эти наблюдения были сделаны позднее, в середине восемнадцатого века (1737), когда Реомюр опубликовал убедительное доказательство партеногенеза. Несколько позднее наблюдения Реомюра заставили Боннэ произвести ряд классических исследований над размножением у тлей, причем работы и рассуждения Боннэ пробудили интерес к партеногенезу — интерес, продолжающийся и до настоящего времени.
Обнаружение Дзирзоном (1845) партеногенеза у пчел вызвало большую дискуссию и прилив нового интереса к этому вопросу. Возникла обширная литература, содержащая наблюдения над другими насекомыми. Распространение теории Дзирзона и применение ее к другим животным, особенно фон-Зибольдом и Лейкартом, связало эту проблему с общими вопросами биологии, в особенности позднее в связи с изучением чисел хромосом в яйцах животных, развивающихся путем естественного партеногенеза. Еще позже опыты с яйцами, искусственно стимулируемыми различными агентами к партеногенетическому развитию, выдвинули другие вопросы, касающиеся роли сперматозоида в оплодотворении.
Важность связи между хромосомами и партеногенетическим развитием была выдвинута на передний план главным образом благодаря наблюдениям и теориям Вейсмана (1886), установившего, что у тех животных, у которых {156} некоторые яйца нормально развиваются без оплодотворения, последние выделяют только одно полярное тельце, тогда как оплодотворенные яйца того же самого животного выделяют два полярных тельца. Эти наблюдения были сделаны несколькими годами раньше, но их значение тогда не было понято. В настоящее время обнаружено, что это явление имеется не только у некоторых ракообразных (Cladocera и Ostracoda), но также и у коловраток, тлей, пчел и муравьев.
Вскоре было установлено, что во многих случаях естественного партеногенеза неоплодотворенные яйца дают либо только самцов, либо только самок. Образование самцов только из неоплодотворенных яиц широко распространено у многих видов пчел и родственных им форм. У других групп, у которых происходит чередование поколений (двуполое поколение чередуется с партеногенетическим), положение вещей более сложно. Прекрасная сводка этого вопроса дана недавно Ванделем. Несколько примеров послужат для иллюстрации основных фактов, известных в настоящее время.
У коловраток, особенно у Hydatina senta, не только выяснено отношение хромосом к половой и партеногенетической фазам жизненного цикла, но проделана большая экспериментальная работа по выяснению причин, ведущих к переходу от одной фазы к другой. Жизненный цикл коловраток состоит из нескольких поколений самок, яйца которых развиваются партеногенетически (девственно), за которыми следуют самцы и самки, размножающиеся половым путем (рис. 89 и 84). Яйца партеногенетических самок выделяют одно полярное тельце. В это время каждая хромосома продольно расщепляется, как при соматическом делении, одна дочерняя половинка направляется в полярное тельце, а другая остается в яйце, которое, следовательно, сохраняет свой полный набор хромосом. Этот способ размножения продолжается в течение нескольких или даже многих следующих друг за другом поколений самок, число которых при неизменности внешних условий неопределенно. Со временем, однако, может появиться самка, образующая яйца различных сортов и называемая самкой, дающей самцов. Судьба яйца такой самки может быть двоякой: во-первых, если самка не была {157} оплодотворена спермой самца, она образует мелкие яйца, выделяющие два полярных тельца; эти яйца развиваются в гаплоидных самцов. Число хромосом при одном из двух делений полярных веретен уменьшается, и яйца сохраняют только половинное число их, которое характеризует также и клетки самца, представляющего собой очень плохо развитое существо, не могущее питаться и внутренность
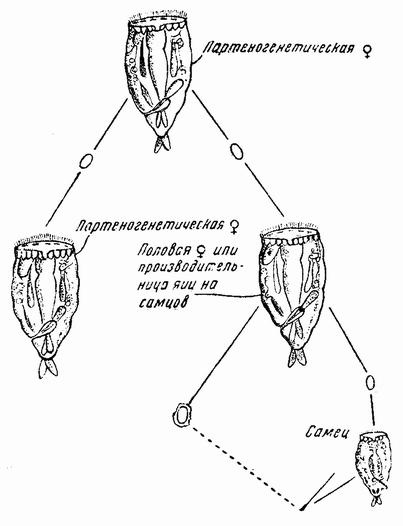 |
|
Рис. 89. Жизненный цикл Hydatina senta. |
которого заполнена главным образом семенными клетками. Во-вторых, если эта же самка была оплодотворена спермой самца вскоре после того как она вылупилась из яйца, то сперматозоиды находят себе путь к молодым яйцевым клеткам и проникают по одному в каждую из них. Эти яйца вырастают тогда до более крупных размеров, чем не оплодотворенные яйца на самцов и покрываются скорлупой. Каждое яйцо выделяет два полярных тельца и во время одного из двух полярных делений число хромосом уменьшается {158} вдвое. Затем ядра яйца и спермия соединяются вместе для образования сегментационного ядра, которое, следовательно, имеет диплоидное число хромосом. Этот эмбрион всегда развивается в самку, которая называется самкой-основательницей и от нее начинается новый партеногенетический цикл.
Существует другой важный факт, касающийся стадий созревания семенных клеток гаплоидного самца. Повидимому, образуются два сорта сперматозоидов, из которых функционирующим является только один из них. Каждая зрелая семенная материнская клетка, являющаяся гаплоидной, делится один раз, причем все хромосомы расщепляются на две половинки, расходящиеся к противоположным полюсам; каждая клетка до сих пор содержит гаплоидное число хромосом. Одни из этих клеток, не претерпевая второго деления, становятся функционирующими сперматозоидами, другие же или некоторые из последних делятся еще раз и дают начало нефункционирующим сперматозоидам. Можно, следовательно, сказать, что единственными функционирующими сперматозоидами являются спермин дающие самок, но имеется ли здесь механизм половых хромосом и если имеется, то как эти хромосомы разделяются, неизвестно.
Одной из наиболее интересных особенностей в жизненном цикле коловраток является то, что внешние условия могут так влиять на партеногенетическую линию, что в ней начинают появляться половые формы. Влияние внешних условий изучалось с разных сторон и, как это было описано выше, было исследовано несколько агентов так называемых производителей-самцов для повышения числа особей, дающих начало самцам. В число этих агентов входят температура, кислород, пища и аккумуляция побочных продуктов обмена по мере старения или перенаселения культуры. Было с убедительностью показано, что из всех этих агентов эффективен только один, а именно пища. Так, например, если Hydatina питается бесцветными жгутиковыми инфузориями (Polytoma), то она существует неопределенно долгое время как партеногенетическая линия; но если в какой-либо момент в культуру будут в изобилии введены зеленые жгутиковые, Chlamydomonas, то почти все следующее поколение самок образует яйца на самцов, а если самки были оплодотворены, то половые яйца. Окончание партеногенетической линии и начало {159} половой фазы обусловлены в этом случае изменением пищи. Как установлено выше, та же самая особь, которая образует яйца на самцов, при раннем оплодотворении, образует также и половые яйца. Можно сказать, что внешняя среда создает пол, но не определяет его. Определяется же пол присутствием или отсутствием сперматозоида.
У одной группы ракообразных, у Cladocera, обнаруживается некоторое сходство с коловратками в том отношении, что у них может быть длинный ряд диплоидных пар-
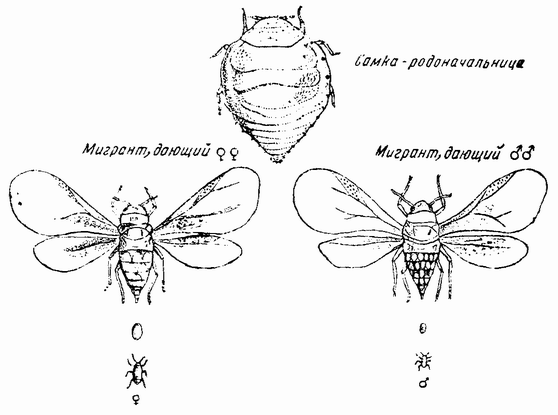 |
|
Рис. 90. Жизненный цикл Phylloxera caryecaulis. |
теногенетических самок, яйца которых выделяют только одно полярное тельце. Как результат изменения во внешних условиях могут появиться самки, дающие начало самцам или половым яйцам, но условия, определяющие тип яйца, в этом случае являются, повидимому, не теми что у коловраток. Например, самки Cladocera, дающие самцов, появляются при перенаселении культуры, а у коловраток, наоборот, перенаселение препятствует появлению таких самок. Предполагалось, что у Cladocera аккумуляция продуктов выделения самих животных (или присутствующих бактерий) вызывает повышенное образование самцов. Прибавление мочевой кислоты, хлористого этана или углекислоты увеличивает процент самцов. Данных о {160} том, что эти агенты вызывают изменения и у коловраток не имеется, наоборот, в застаревших культурах на конском навозе процент самок производящих самцов уменьшается.
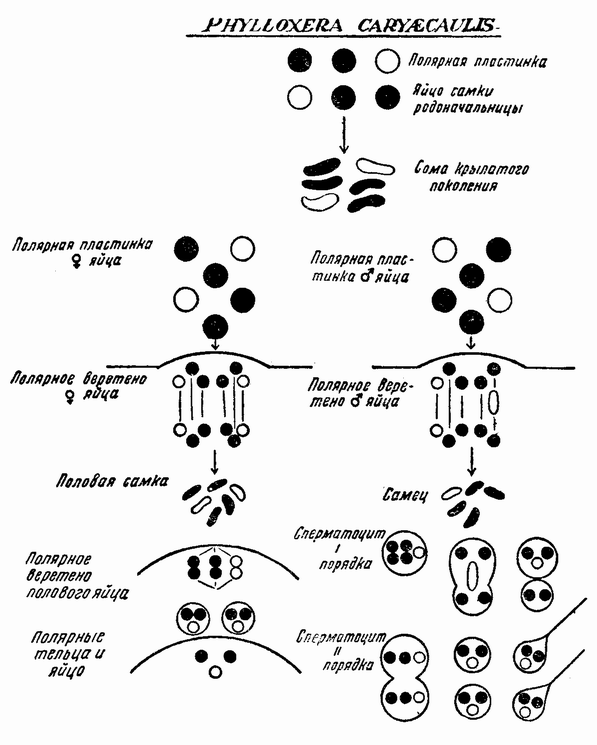 |
|
Рис. 91. Хромосомный цикл P. caryecaulis. |
У насекомых чередование партеногенетического и полового поколений имеют также тли и филоксеры (рис. 90). У первых было показано, что длина дня, т. е. светлых часов, является одним из агентов, вызывающих изменение, но {161} неясно, действует ли этот фактор непосредственно или каким-то косвенным путем. В этих двух группах известны два других факта, объясняющих некоторые отношения, не обнаруженные у коловраток (l). Партеногенетические самки имеют диплоидное число хромосом и выделяют одно полярное тельце (рис. 91). В конце партеногенетической линии появляется самка, яйца которой тоже выделяют только одно полярное тельце, но, когда все другие хромосомы делятся, одна из Х-хромосом целиком отходит в полярное тельце, оставляя своего партнера — другую половую хромосому — позади (рис. 91, справа). Это яйцо, следовательно, дает начало самцу, который имеет диплоидное число хромосом минус один Х(2). Когда семенные клетки у этого самца претерпевают первое деление созревания, то каждая из хромосом коньюгирует со своим партнером за исключением лишь Х-хромосомы, которая не имеет партнера. Члены каждой пары движутся затем к противоположным полюсам, а единственная Х-хромосома к одному из полюсов (рис. 91). Клетка, получающая Х-хромосому, становится крупнее другой и две дочерних клетки разделяются. Одна из них имеет гаплоидный набор (включая X), другая же содержит гаплоидный набор минус Х-хромосому. Крупная клетка претерпевает затем второе деление созревания, при котором каждая хромосома, также как и Х-хромосома, расщепляется. Каждая клетка становится функционирующим спермием на самку, так как она содержит Х-хромосому. Меньшая же клетка, являющаяся продуктом первого деления, дегенерирует.
У других самок, появляющихся одновременно или на поколение позже, чем самки, дающие самцов, развиваются половые яйца. Они выделяют два полярных тельца и сохраняют гаплоидное число хромосом, в числе которых имеется одна Х-хромосома (рис. 91, слева). При оплодотворении спермием на самку, восстанавливается полное число хромосом (2А+2Х). Из этого яйца, называемого покоящимся яйцом, развивается самка-основательница, являющаяся родоначальницей новой линии партеногенетических самок.
Здесь половые хромосомы опять играют свою обычную роль в определении пола, причем фактором, изменяющим партеногенетическую фазу в половую, является внешняя среда.
Существуют различные виды животных, представленные в некоторых географических областях только самками, {162} а в других и самками и самцами. Первые размножаются путем партеногенеза; вторые — из оплодотворенных яиц. Например: один из равноногих раков, Trichoniscus, представлен на севере Европы только самками, которые являются триплоидными и содержат 24 хромосомы; на юге же найдены диплоидные самцы и самки, имеющие 16 хромосом. Если яйца этих половых самок не будут оплодотворены, то они не развиваются. Здесь, следовательно, имеется две определенных расы одного и того же вида, причем, так как они не могут скрещиваться между собой, можно сказать, что это два вида, которые идентичны во всем, кроме способа их размножения.
У другого ракообразного, Arthemia salina, имеются тетраплоидные партеногенетические расы с 84 хромосомами и диплоидные половые расы с 42 хромосомами. Среди кузнечиков известны также случаи, когда яйца при оплодотворении дают начало и самцам и самкам, причем пол особей регулируется двумя сортами сперматозоидов, а без оплодотворения образуют только самок. Некоторые виды Phasma размножаются только партеногенетически; другие же виды имеют половое размножение. Можно было бы привести много других случаев, в которых партеногенез занимает определенное место в жизненном цикле организма.
Эти примеры, число которых может быть увеличено, служат для демонстрации распространенности партеногенеза в животном царстве. Однако это явление известно также и у многих растений. Эти случаи подчеркивают тот факт, что способность к развитию заключается в самих яйцах. Получается, следовательно, картина, отличающаяся от обычного представления, подчеркивающего, что начало развития является результатом оплодотворения. Я думаю, что правильнее будет сказать, что главная функция проникновения спермия в яйцо заключается в удалении препятствия, удерживающего яйцо от развития. С этой точки зрения партеногенетическое яйцо отличается от обычного тем, что это препятствие в нем отсутствует. Конечно, имеют значение и другие обстоятельства, в частности число содержащихся в яйце хромосом, число выделяемых полярных телец и роль сперматозоида, заключающаяся либо в введении в яйцо центра деления, либо в вызывании там его развития. Эти взаимоотношения станут более ясными, когда будет описано поведение яиц, искусственно стимулируемых к развитию. {163}
Еще в 1866 году Тихомировым было обнаружено, что неоплодотворенные яйца шелковичной бабочки при трении между двух кусков ткани или при погружении на короткое время в серную кислоту начинают развиваться (6%) и образуют зародыши. Позднее этот опыт был повторен различными путями и в целом подтвержден, но была лишь сделана оговорка, что даже и без воздействия неоплодотворенные яйца некоторых рас этой бабочки тоже могут начать развиваться. Примерно тридцатью годами позднее было обнаружено, что неоплодотворенные яйца морского ежа и одной аннелиды можно заставить развиваться по крайней мере до дробления путем прибавления к морской воде некоторых солей. Развития неоплодотворенных яиц морского ежа в плавающий зародыш не удавалось получить до 1900 года, когда Лёбу удалось вызвать это путем изменения химического состава морской воды. С тех пор было показано, что яйца можно заставить развиваться многими различными агентами. Нельзя сказать, чтобы эти опыты осуществили давнишнюю надежду, что «химическое оплодотворение» разъяснит природу нормального оплодотворения, но рассмотрение изменений, вызываемых в яйце искусственными агентами, привело к некоторым интересным рассуждениям относительно природы изменений, которые может совершать сперматозоид при проникновении в яйцо. Трудности интерпретации этой проблемы обусловлены не отсутствием фактов, а открытием весьма многих путей, по которым может начаться развитие. Установить какое-либо общее свойство для всех этих агентов оказалось невозможным. Все попытки объяснения сбивались до сих пор с толку изменениями в самом яйце. Это станет очевидным, когда будут рассмотрены некоторые из применявшихся методов.
В одном из наиболее ранних опытов яйца морского ежа помещались примерно на час в морскую воду, к которой был добавлен хлористый магний (50 см3 морской воды (50 см3 2,5-нормального раствора MgCl2), после чего они переносились обратно в морскую воду. Более совершенный метод состоял в переносе яиц на два часа в гипертоническую морскую воду, а затем в воду, содержащую немного уксуснокислого этила. При возвращении в морскую воду многие яйца развиваются нормальным образом. Подобный же результат дают также некоторые кислоты. {164} Первые указания на то, что яйцо будет развиваться, заключаются в отставании его оболочки. Это происходит также и при нормальном прободении яйца спермием. Появляется прозрачная оболочка, которая отстает от яйца и довольно скоро отделяется от поверхности его, оставляя внутри пространство, заполненное жидкостью. Раньше предполагалось, что одним из существенных факторов в начале развития яйца является образование оболочки, но с тех пор были найдены другие способы воздействия, препятствующие отставанию оболочки и тем не менее не влияющие на нормальное развитие.
Яйца морской звезды можно заставить развиваться, воздействуя на них в определенный момент морской водой, насыщенной углекислотой. Дробление яиц вызывается даже встряхиванием или воздействием масляной кислотой. Яйцо лягушки можно заставить развиваться путем накалывания его тонкой иголкой. Важно ли или нет для успеха наличие на иголке следов лимфы или крови не ясно, но некоторые опыты, поставленные для проверки значения этого момента, показывают, поводимому, что хотя в отсутствии лимфы развитие и может начаться, но фигура деления при этом не развивается, в то время как в присутствии лимфы некоторые яйца могут делиться. Из тысяч наколотых яиц в лучшем случае только одно может дать начало молодому зародышу, и очень редко случается, что какой-либо из них развивается до стадии лягушки. Яйца некоторых аннелид могут быть стимулированы к развитию хлористым калием и натрием, яйца же других — кислотами, температурой или, наконец, разбавлением морской воды.
На основании этих данных очевидно, что хотя и имеются специфические агенты, дающие наилучший результат для каждого рода яиц, но эти агенты настолько различны, что невозможно сделать общее заключение о способе их действия. Лёб придавал большое значение цитолитическому эффекту этих агентов на поверхностный слой яйца. В известном смысле эффект этот можно назвать разрушительным. Лёб предполагал, что двойное воздействие дает лучший результат, потому что второе воздействие противодействует эффекту цитолитического действия первого. Но так как во многих случаях достаточно единственного воздействия, то из этого не следует, что этот эффект является цитолитическим, а даже если это и {165} так, то природа такого изменения еще не выяснена. Необходимо выяснить, каким образом эти агенты удаляют препятствие, подавляющее изменения, которые готово претерпеть само яйцо. Несомненно, что некоторые из применявшихся агентов действовали на яйцо повреждающим образом, ибо при оставлении в этих жидкостях дольше оптимального времени, яйца безвозвратно повреждались. Начальным изменением на поверхности яйца может быть
 |
|
Рис. 92. а, b, с — срезы через яйцо лягушки, стимулированное к развитию накалыванием чистой иголкой, видно образование звезды около места прокола; d, е, — срезы через яйца, наколотые иглой с кровью (двойное воздействие) (по Герланту). |
шаг втом же самом направлении, но термин цитолитический процесс недостаточно объясняет, что именно при этом происходит; разве только это может означать какого-то рода изменение, повреждающее какую-нибудь структуру на поверхности яйца или контролирующее его развитие.
В последние годы много внимания было уделено тем изменениям, которые могут быть видимыми в самом яйце после воздействия на него активирующими факторами. Это главным образом касается развития в протоплазме центров деления и поведения яйцевого ядра. Короче {166} говоря, было найдено, что все успешные опыты приводили к развитию в яйце звезд и что лучшими воздействиями были те, при которых вблизи ядра появлялось немного звезд, предпочтительно только две. Между звездами развивается веретено, к которому прикрепляются хромосомы, и затем может произойти деление. Надо отметить, что в нормально оплодотворенном яйце звезда появляется вблизи проникающего в яйцо ядра спермия и делится для образования полюсов веретена. Звезда, лежащая на внутреннем конце веретена второго полярного тельца, к этому моменту исчезает. Доказательства восстановления или сохранения этой звезды в яйце, искусственно стимулированном к развитию, отсутствуют, наоборот, как установлено выше, звезда появляется de novo. Более того, фигура деления не образуется путем деления одной из этих звезд, а возникает из одной или большего числа их, возникших независимо. Создается впечатление, что хромосомы посредством их прикрепляющих нитей находят эти веретена и используют их.
То, что происходит с ядром, зависит, повидимому, как от рода яиц, так и от характера воздействия. В зрелом яйце морского ежа, где образование полярного тельца закончилось, ситуация относительно проста. Гаплоидное ядро делится в нем, и каждая получающаяся клетка имеет гаплоидное же число хромосом. Клетки личинки, повидимому, тоже гаплоидны. Возможно, что случайно хромосомы могут разделиться, в то время как деления яйца не происходит. Таким образом, яйцо становится диплоидным прежде, чем происходит следующее (которое теперь является первым полным) деление и образуется диплоидный зародыш. Кажется вероятным, что те редкие случаи, в которых зародыш развивался до взрослой формы, являются как раз теми, в которых такая диплоидность имела место.
Практически тот же самый вывод применим и к яйцу лягушки. После накалывания в нем выделяются два полярных тельца, и гаплоидное яйцевое ядро отходит к центру яйца, где оно входит в соприкосновение с двумя появившимися в яйце звездами (рис. 92). Гаплоидные эмбрионы выглядят слабыми. В начале развития количество протоплазмы у них вдвое превышает то, которое соответствует числу их хромосом. Не ясно, является ли именно это первичной причиной их слабости, и неизвестно, происходит ли на более поздних стадиях регуляция между гаплоидным {167} числом хромосом и величиной клетки. Все особи, развившиеся до взрослых стадий, оказались диплоидными. Знание поведения хромосом при естественном партеногенезе делает возможным изучение вопросов наследственности. Так как в этих случаях выделяется только единственное полярное тельце и хромосомы продольно расщепляются, как и в обычном митозе, то все потомство женской линии имеет в точности одинаковое генетическое строение. Благодаря генетической идентичности этих самок, вся индивидуальная изменчивость, которую они обнаруживают, может быть отнесена за счет небольших различий во влияниях среды. Проведенные измерения действительно, обнаружили в этих случаях такое же близкое сходство, какое имеется у идентичных близнецов, причем это происходит здесь по той же самой причине, ибо в обоих случаях особи имеют идентичные гены. Наоборот, внезапные и часто большие как структурные, так и физиологические изменения, которые имеют место при окончании партеногенетического цикла и при начале половой фазы дают возможность изучить, в какой степени некоторые новые внешние влияния смогут дать новый конечный результат на том же самом генетическом фоне. Здесь опять возникает уже обсуждавшийся ранее вопрос, а именно, обусловливается ли изменение началом действия различных генов, или же продукт всех генов остается тем же, а внешняя среда заставляет эти продукты давать отличный конечный результат. Ответ и в этом случае не убедителен, но так как было показано, что изменение происходит внезапно, действуя на организм скорее как стимул, а не продолжительно в течение всего развития, то может казаться более вероятным, что на критической стадии приходят в действие различные гены.
| << | {168} | >> |
Иногда сравнивают развитие целого эмбриона из фрагмента яйца с восстановлением потерянной части взрослого животного. Если бы такое сравнение имело какое-нибудь основание, то неспособность некоторых изолированных бластомеров создать целый эмбрион следовало бы сравнивать с неспособностью некоторых взрослых животных заменять потерянные части. В действительности же такие сравнения не очень поучительны. Но тем не менее регенеративные процессы у животных содержат в себе некоторые необычайно интересные факты, имеющие отношение к закономерностям индивидуального развития. В восстановлении целого эмбриона из части яйца могут участвовать частично те же самые факторы, что и в реставрации целого простейшего или целой гидры из их частей.
Существует два главных типа регенерации. При одном из них новая структура развивается путем реорганизации старых материалов; при другом новые структуры формируются из новых материалов, образующихся из старой части.
Прекрасный пример первого случая обнаружен у простейшего — Stentor (рис. 93 А). Если стентора разрезать на две части (рис. 93 А1, а, b), то каждая часть округляется и в течение дня, или даже скорее, реорганизуется в стентора половинной величины, у которого, впрочем, на время остаются некоторые признаки той части, из которой этот участок произошел. Это лучше всего видно, когда животное сокращается и вытягивается (рис. 93 а1, b1).
Другой пример прямой трансформации части в новый организм обнаружен у многоклеточного животного — гидры (рис. 94 А). Если гидру разрезать на несколько частей, то каждая часть сокращается, закрывает свои открытые концы и через несколько дней постепенно принимает {169} типичную форму этого животного (рис. 94 а—e). Около переднего конца вырастают небольшие щупальца, отрезок становится длиннее и тоньше и на одном конце его образуется рост, а на другом подошва.
Родственное гидре животное, гидроид Tubularia, тоже регенерирует из старых тканей, но так как оно заключено в жесткую хитиновую оболочку, то отрезанная частица
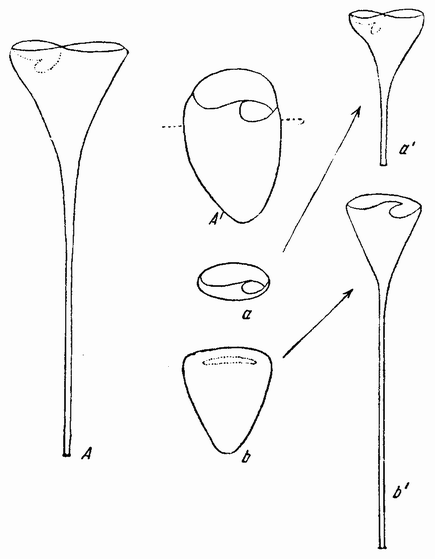 |
|
Рис. 93. Stentor coeruleus в вытянутом (А) и в сократившемся (А1) состоянии. На рис. а и b показано место разреза на две части; а1 и b1 — регенерация из каждой части двух новых стенторов (по Моргану). |
не сокращается и не перестраивается сначала в новое целое меньших размеров, а новый гидрант с самого начала оказывается почти такой же величины, как и исходный. Tubularia является колониальной формой, состоящей из ветвящегося в основании ствола, образующего трубчатые стебельки, каждый из которых заканчивается головкой {170} или гидрантом, имеющим два ряда щупалец, перистом и рот (рис. 95 а). Если гидрант разрезан, то открытый конец стебелька менее чем в полчаса покрывается слоем клеток, которые выталкиваются из поверхности разреза стебелька. Покрывающие клетки являются не новыми, а представляют собой старые клетки из окружающей разрез стенки. Приблизительно через 12 часов около срезанного конца
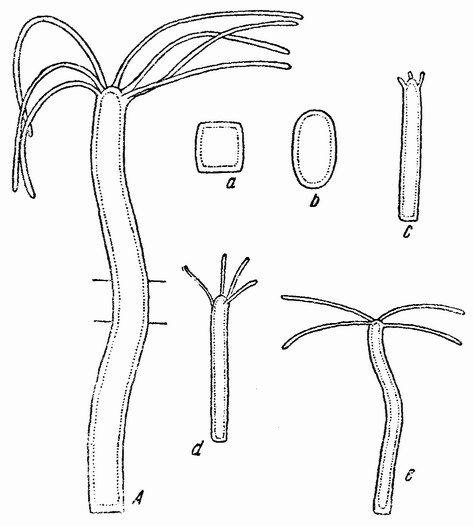 |
|
Рис. 94. А — Hydra viridis; a — кусочек, вырезанный из ее середины и его развитие (b, c, d, е) (по Моргану). |
появляются два ряда пигментированных полосок (рис. 95 b), из которых в дальнейшем развиваются два ряда щупалец. Эти полоски отделяются от поверхности по всей своей длине, прикрепляясь к стебельку только своим основанием, и становятся щупальцами. Над внешним кругом щупалец образуется перистом с новым ртом на его конце. Затем новый гидрант выдвигается вперед за срезанный конец хитиновой трубки (рис. 95 с). Гистологическое {171} изучение такой части в то время, когда происходят эти изменения, показало, что новых клеток при этом либо вовсе не образуется, либо их образуется очень мало. В новые части, шупальцы, перистом и т. д., непосредственно превращаются старые клетки.
Стенки стебелька Tubularia состоят из двух слоев клеток, располагающихся с внутренней стороны кутикулы. Широкий центральный пищевод неполностью разделяется продольной перегородкой, по одной стороне ко-
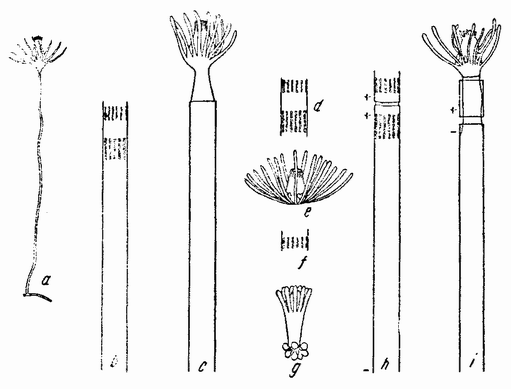 |
|
Рис. 95. Регенерация у Tubularia. а — гидрант и стебелек; b — появление вблизи срезанного конца двух кругов щупалец; с — гидрант, развившийся из b, d, е — регенерация гидранта без стебелька, из короткого кусочка; f, g — регенерация только дистального конца гидранта; h — короткий кусочек, привитой на конец длинной части в обратном положении; i — регенерация из него единого гидранта (по Пиблсу). |
торой пищеварительная жидкость циркулирует вверх, а по другой вниз, проходя вокруг свободного конца перегородки как раз под гидрантом. Вскоре после удаления старого гидранта перегородка поблизости от поверхности среза разрушается, и ее клетки, содержащие зерна красного пигмента, попадают в пищеварительную жидкость. Так как при появлении щупальцевых полосок в них развивается красный пигмент, то до того, как его происхождение стало известным, предполагалось, что красный пигмент пищеварительной жидкости является формообразовательным {172} веществом. Однако этот взгляд упускает из вида тот простой факт, что во время закладки пигментированных полосок зерна пигмента циркулируют по всей длине стебля, а не собираются на его переднем конце. Только после закладки щупалец и образования головы зернышки слипаются вместе в желудке нового гидранта и извергаются изо рта, когда гидрант выдвигается вперед над старой кутикулярной стенкой. Попытка усмотреть в этом случае формообразующее вещество была явно преждевременна.
В качестве примера регенерации путем развития новых клеток может быть выбран процесс, происходящий у пресноводного кольчатого червя Lumbriculus. Если этот червь разрезан на две части, то новый хвост вырастает из заднего конца передней части, а новая голова из переднего конца задней части. Через несколько дней после разреза на срезанной поверхности появляется нарост из клеток, нг обнаруживающих диференцировки старых клеток, из которых они произошли. Они стали более округлыми и потеряли в некоторой степени их видимые морфологические черты. В деталях эти изменения заключаются в следующем: срезанный конец закрывается стягиванием стенок тела. Затем, когда стенки расслабляются, слой эктодермальных клеток наползает на конец, некоторые из этих клеток втягиваются и начинают заполнять собой внутренность. Далее эти внутренние клетки начинают диференцироваться заново. Из внутренних клеток нароста происходят мозг, соединительная ткань и брюшная цепочка формирующегося головного участка, а также мускулатура. На конце вдается внутрь трубка, образованная из поверхностной эктодермы и из которой развивается пищевод, внутренний конец которого соединяется с передним концом пищеварительного тракта и открывается в него. По мере удлинения нового нароста он обнаруживает циклические перетяжки на шесть или семь колец, или сомитов. Затем процесс заканчивается, причем замещения всего переднего конца не происходит (если было удалено больше чем семь или восемь сегментов), а восстанавливается только часть его, достаточная для образования головы.
Образование нового хвоста происходит сходным образом за исключением лишь того, что после образования нескольких первых сегментов на конце образуется растущая зона, которая прогрессивно добавляет новые сегменты {173} к образованным в начале и которая в конце концов может дать начало отрезанному числу сегментов.
Самая важная особенность регенерации новых частей у этого червя заключается в происхождении и судьбе новых клеток. Новый кожный покров происходит из клеток старого покрова, причем это относится также и к новому мозгу, нервной цепочке и даже к новой мускулатуре. Насколько известно, все эти три ткани или органа происходят из одних и тех же исходных клеток, которые раньше выполняли функции кожного покрова червя. Какая из трех возможностей реализуется, будет, повидимому, определяться локализацией или положением этих клеток в новой части. Каким из этих фактов придать большее значение, долгое время являлось загадкой для исследователей. Одно время считалось, что сходные ткани образуют сходные, но очевидно, что это верно только частично: для Tubularia это так, но у Lumbriculus это верно только для кожного покрова. Предполагалось также, что регенерирующие органы образуются таким же образом, как и в эмбрионе, т. е. из тех же зародышевых листков, однако, для мускулатуры у Lumbriculus это не верно.
Это еще раз выдвигает вопрос о роли генов в этих регенеративных изменениях. Так как все клетки содержат полный комплект генов, то, казалось бы, каждая клетка способна к образованию любой части организма, если только ее протоплазма не была уже раньше необратимо изменена в каком-нибудь определенном направлении. Исходя из этого предположения, должно казаться более вероятным, что старые клетки будут продолжать функционировать в новой части так же, как и раньше, и во многих случаях это, очевидно, соответствует действительности; но к клеткам, которые превращаются в другие ткани, это объяснение уже не будет применимо. Можно, пожалуй, сказать, что после того, как клетки теряют связь с тканями, из которых они происходят, некоторые из них могут утерять свою диференцировку (и наблюдения подтверждают это) и начинают развиваться согласно своему местоположению. В чем заключается значение «местоположения», трудно сказать, но путь для решения этого вопроса все же имеется. Новая диференцировка, по крайней мере в некоторых случаях, начинается в точке соприкосновения с той из старых тканей, которая лежит ближе всего к ней. Старая ткань действует на новую как диференциатор или организатор, {174} и мы можем, пожалуй, предположить, что это влияние является химическим. Однако проводить такое сравнение слишком далеко рискованно. Этого рода данные не помогают решить вопрос, действуют ли все гены в то время, когда в клетках происходят эти изменения, или же некоторые гены вызываются к действию новой средой, окружающей клетки в новом участке.
Как было установлено в главе I, одной из первых попыток объяснить не только как эмбриональное развитие, но и процессы регенерации является гипотеза Ру-Вейсмана, которая покоится на предположении, что при каждом дроблении яйца происходит качественное разделение веществ заключенных в хромосомах. Дочерние клетки приобретают, следовательно, различное проспективное значение. Как уже говорилось, это предположение не было в соответствии с известными уже в то время наблюдениями, что хромосомы в точности расщепляются пополам, и еще более не гармонирует с современными представлениями о генной структуре хромосом. Вейсман пытался сопоставить эту же гипотезу с фактами регенерации. Так как новая часть, развивающаяся из любого места разреза, подобна удаленной части, то он предполагал» что в каждом районе организма, после того как произошла окончательная диференцировка, остаются остаточные клетки, «назначением» которых является замещение дистальных по отношению к ним частей. Иными словами, при развитии организма используются не все клетки определенного сорта, а некоторые из них остаются в тканях, и именно из них и развивается новая часть. Эта гипотеза до некоторой степени объясняла бы также, почему сходные ткани образуют сходные с ними. В действительности же имеется большое число фактов, которые не находятся в соответствии с этим взглядом, ибо клетки, образующие новую часть, могут возникать из части тела, совсем отличной от той, которая дала начало рассматриваемому эмбриональному органу.
Прекрасный пример последней точки зрения найден в регенерации хрусталика в глазу саламандры (рис. 96). Если хрусталик удален, то новый хрусталик развивается из края радужницы. У эмбриона хрусталик образуется из внутреннего слоя покрывающей эктодермы. Следовательно, регенерировавший хрусталик развился из части тела, имеющей эмбриональную историю, совсем отличную от истории первоначального хрусталика. Более того, он {175} развился из функциональных клеток старой радужницы, а не из остаточных клеток. Можно сказать, что новый хрусталик развился из ткани, которая имеет наилучшее из возможных положений для образования такого хрусталика. С другой стороны, большинство клеток на краю радужницы содержит пигмент, тогда как клетки хрусталика прозрачны.
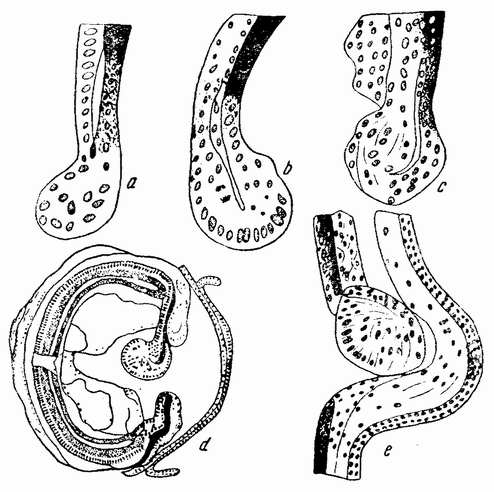 |
|
Рис. 96. Регенерация глазного хрусталика у тритона после удаления старого хрусталика; a—e — стадии развития нового хрусталика, образующегося на краю радужницы; d — то же самое, видно положение нового хрусталика (по Вольфу и Фишер). |
В этом случае может казаться, что если имеются особые гены, активность которых объясняет сохранение пигментных клеток радужницы, то должны быть другие гены, которые становятся более активными, когда хрусталик теряет связь с радужницей. Но результат может быть одинаково хорошо объяснен, исходя из предположения, что в обоих случаях все гены активны и что регенерировавший орган первично детерминирован новой средой,в которой находятся клетки края радужницы. Протоплазма здесь может, следовательно, считаться отличной от той, которая {176} была раньше, вследствие чего при том же самом продукте генов конечный результат является тоже отличным. Конечно, это заключение не объясняет специфической природы реакции. А это как раз и представляет собой суть спора. Неудивительно поэтому, что, благодаря нашей неспособности установить, какие факторы участвуют в вызывании этих результатов, ряд эмбриологов прибег к сверхъестественным, т. е. метафизическим, попыткам их объяснения. Наиболее популярное из этих объяснений известно под именем витализма, причем в отдельных обсуждавшихся случаях в качестве виталистического агента призывалась энтелехия. Этот взгляд, пожалуй, даже не заслуживает критики, поскольку он основывается на метафизической концепции, но по крайней мере некоторые замечания следует все-таки сделать: во-первых, такое объяснение также трудно «понять», как и объяснить с его помощью факты; во-вторых, если с ним согласиться, то, благодаря допущению целесообразности, оно будет тормозить все дальнейшие попытки найти натуралистическое объяснение; и, в-третьих, при изучении регенерации получены некоторые факты, ставящие это учение в очень смешное положение, поскольку энтелехия выступает как благотворительный фактор.
Например, если дождевой червь разрезан по середине на две части, то на заднем участке развивается голова, состоящая из четырех или пяти сегментов, но так как область, содержащая репродуктивные органы, не регенерирует, то такое «животное» никогда не может размножиться. Если червь разрезан ближе к заднему концу, то на переднем конце задней части (рис. 97 d, е) развивается хвост, который продолжает прибавлять новые сегменты, но такая часть с двумя хвостами через некоторое время погибает, так как она не может принимать пищу.
Подобный же результат получается, когда у планарии отрезается небольшой кусочек на конце хвоста. В этом случае на переднем конце отрезанной части регенерирует другой хвост. Если у планарии отрезан конец головы, то на задней поверхности среза развивается новая голова (рис. 91 а, b, с). Двухголовая часть погибает, так как у ней не образуется рот. Получается, таким образом, что энтелехия как будто бы может делать грубые ошибки, при чем при этих условиях это происходит постоянно, что и поощряет поиски другого объяснения. {177}
У высших растений большинство регенеративных явлений настолько отлично от регенерации животных, что сравнивать их трудно. Удаление терминального конуса роста сопровождается развитием нескольких преформированных почек, которые уже присутствуют ниже по стеблю. Здесь вопрос заключается скорее в агенте, который контролирует развитие почек, причем имеются данные, доказывающие, что это может быть связано с появлением какого-то веще-
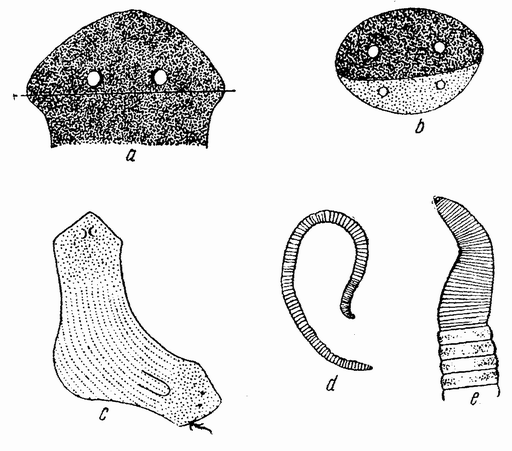 |
|
Рис. 97. а — голова Planaria lugubris, видно место разреза; b — регенерация новой головы на заднем конце старой головы; с — более крупный кусочек с глоткой и двумя головами; d — хвост дождевого червя, у которого на переднем срезе регенерировал хвост; е — то же самое, увеличено (по Моргану). |
ства, вырабатывающегося в верхушке стебля. Субстанция, представляющая собой это вещество, была выделена и ее действие показано экспериментально. Следовательно, химическое вещество имеет по крайней мере некоторые свойства энтелехии.
Другие растения могут регенерировать также путем образования маленьких отростков на поверхности среза. Поверхность разреза сначала покрывается неспециализированными клетками из так называемого камбиального слоя. В одной или нескольких точках новой поверхности {178} появляется небольшая почка, которая в одних случаях, вероятно, часто происходит из единственной клетки, в других же — из нескольких клеток. Один из этих отростков может вырасти в терминальный стебель или ветку или же, если он удален и посажен в землю, то он образует корни и станет целым растением.
В двух случаях, выбранных для иллюстрации регенерации у животных путем образования новой ткани, развитие происходило из передней и задней поверхностей раз-
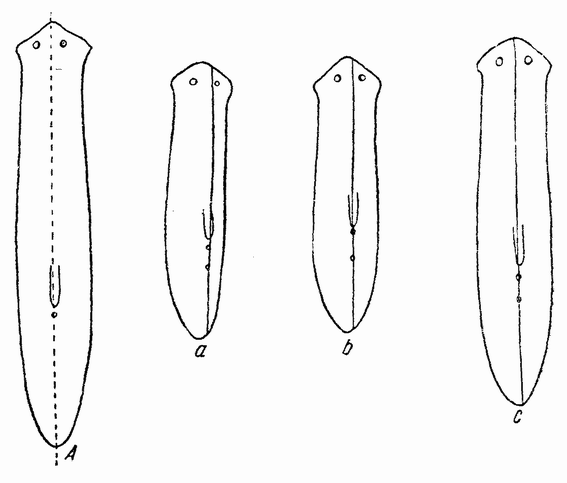 |
|
Рис. 98. А — Planaria, продольно разрезанная на две части; а, b, с — регенерация целого червя из левой половины А (по Моргану). |
реза. Однако может также иметь место и латеральная регенерация, как это, например, происходит, если отрезать ногу саламандры в любом месте. Из оставшейся части конечности регенерирует ровно столько, сколько было отрезано, а отрезанный кусок погибает от недостатка питания. Если отрезана правая нога, то новая нога оказывается правой; если отрезана левая нога — новая нога будет тоже левой. Если плоский червь продольно разрезан надвое (рис. 98 А), то каждая половинка восстанавливает {179} недостающую половину. Левая половина червя (рис. 98 a) образует новую правую сторону и между старой и новой частями устанавливается срединная плоскость (рис. 98 с). То, что происходит в этом случае, подобно тому, что имеет место при разъединении первых двух бластомеров ланцетника. Если бы оба бластомера остались вместе, то один из них должен был бы образовать правую сторону тела, но после изоляции из его вещества развиваются и правая и левая стороны эмбриона, причем это достигается не путем регенерации клеток для отсутствующей половины, а путем установления в сферической бластуле новой срединной плоскости, которая была, так сказать, создана клетками правой стороны. Это же имеет место также и у Stentor, если его разрезать в продольной плоскости на две части.
Другой вопрос, который следует поставить, касается связи между скоростью роста и местом, на котором произведен разрез. Если отрезать кончики хвостов двух дождевых червей, один недалеко от конца, а другой дальше кпереди, то скорость, с которой происходит регенерация, медленнее, когда разрез расположен ближе к концу, и быстрее, когда он находится дальше кпереди. Два задних конца могут восстановить недостающие части в течение одного и того же времени, благодаря более быстрому росту от разреза, расположенного дальше кпереди. Телеологически это можно объяснить либо допущением, что чем большая необходимость в органе испытывается организмом, тем быстрее происходит рост, либо же, что энтелехия действует быстрее в передней половине, потому что там «чувствуется» большая нужда для приведения червя в порядок. Имеется, однако, биологический предел даже и для «мудрости» энтелехии, ибо если разрез сделан еще дальше кпереди, то рост заднего конца вообще не происходит.
То же самое справедливо и для конечности саламандры. Если отрезан конец одного пальца, то его восстановление будет продолжаться столько же времени, как и восстановление всей ноги, отрезанной в ее основании. Очевидно, что нужда в целой ноге больше нужды в кончике одного пальца и т. д. Оставляя в стороне метафизические толкования, факты относительно скорости регенерации в зависимости от места разреза дают материал для некоторых интересных сравнений. Клетки верхушки пальца нормальной, не {180} поврежденной саламандры или конца хвоста нормального червя прекращают размножаться, за исключением лишь размножения клеток для восстановления нормального износа частей. Можно думать поэтому, что каковы бы ни были факторы, ограничивающие максимальную возможную скорость роста в неповрежденном животном, те же самые факторы регулируют скорость роста клеток при регенерации. Неясно, однако, почему ближе к переднему концу скорость регенеративного роста должна быть больше, чем дальше от него. Этот факт требует другого объяснения.
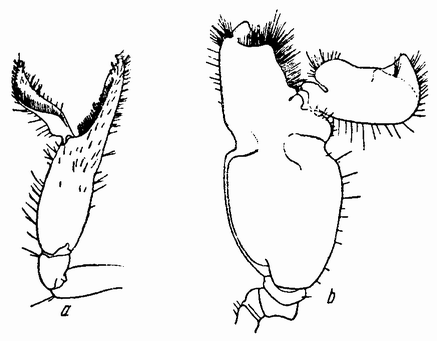 |
|
Рис. 99. Alpheus. а — малая клешня; b — большая клешня (по Вильсону). |
Хорошо известно, что и при голодании кольчатые черви и планарии регенерируют. Старая, голодающая часть активно уменьшается по мере увеличения размера новой части. Но величина новой части у голодающего животного оказывается заметно меньшей, чем у питающегося. Это взаимоотношение подчеркивает тот факт, что общее количество пищи, пригодной для данной части тела, не является фактором, имеющим значение для регенерации, и сказывается только на размерах новой части. Вопрос оказывается более сложным, ибо, кроме скорости роста, еще большее значение имеет характер этого роста, а его отношение к старой части явно зависит не от количества пригодной пищи, а от других обстоятельств.
Нечто в том же роде наблюдается при компенсаторной {181} регенерации, которая может быть лучше всего иллюстрирована на регенерации большой и малой клешни ракообразного Alpheus, у которого обе клешни отличаются не только
 |
|
Рис. 100. А — молодой самец краба на ранней стадии развития. В это время обе клешни более похожи на мужскую клешню; В — молодой краб, потерявший левую клешню, у которого с правой стороны регенерировала новая клешня, похожая на клешню самки (по Моргану). |
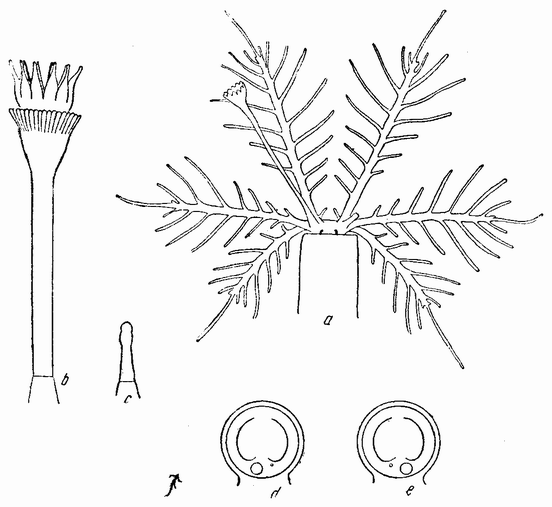 |
|
Рис. 101. а — молодая стадия трубчатого червя; видно, что одна жаберная ветка изменена в крышечку; b и с — функционирующая и рудиментарная крышечка взрослого гидроида; d и е — схемы, показывающие прикрепление этой крышки (по Зеленому). |
величиной, но и формой (рис. 99). Если удалена большая клешня, то в основании ее начинает регенерировать новая. Когда животное линяет, вылезая из своей старой кутикулы, {182} то первоначальная маленькая клешня превращается в большую клешню, а регенерировавшая клешня в меньшую. Если, наоборот, удалена маленькая клешня, то новая оказывается тоже малой, а старая после линьки тоже попрежнему остается большой клешней. Повидимому, здесь имеется своего рода физиологическое равновесие между клешнями обоих сортов, природа которого, однако, неизвестна.
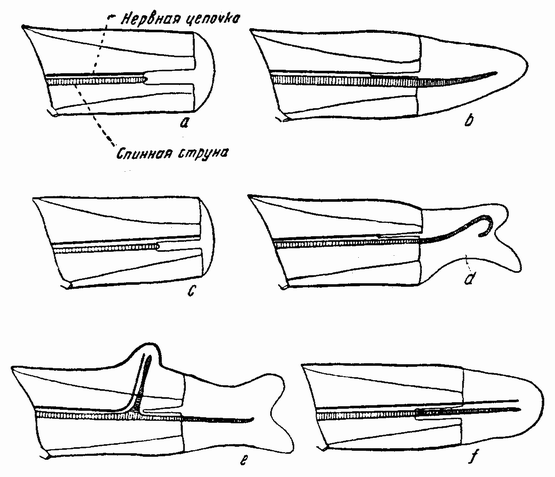 |
|
Рис. 102. Хвост головастика, а — спинная струна и нервный ствол из разреза удалены; с — удалена только спинная струна (нервный ствол остался); b, d, e — хвост регенерирует, когда спинная струна достигает поверхности разреза, но не регенерирует, когда до поверхности разреза доходит только нервный ствол (с) (по Моргану и Дэвису). |
У других десятиногих раков, у которых тоже имеется различие в клешнях обеих сторон, такой перемены при регенерации не происходит. Как было показано, происхождение асимметрии у одного из них, у краба-отшельника, зависит от взаимоотношений, подобных таковым у Alpheus, но в этом случае не происходит реверсии на взрослой стадии. У молодого краба обе клешни сначала одинаковы {183} (рис. 100). Если одна из них теряется, то при следующей линьке она принимает форму малой клешни, после чего реверсия уже невозможна.
У сидячих кольчатых червей имеется пробка или крышечка (рис. 101 b), которая закрывает отверстие трубки, когда животное втягивается в нее. На противоположной стороне имеется рудиментарная крышечка (рис. 101 с). Если большая функционирующая крышечка удалена, то меньшая становится большей. Регенерировавшая крышечка остается малой. Реверсия может быть получена вторично удалением вновь регенерировавшей большей
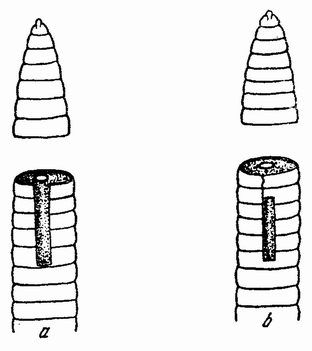 |
|
Рис. 103. а — операция удаления из места разреза вентральной нервной струны; b — то же самое другим путем. Новая голова развивается только в том случае, если на поверхности разреза имеется нервная струна или кусочек ее (b) (по Моргану). |
крышки. Таким образом, в этом случае условия являются такими же, как и у Alpheus.
Наконец, было обнаружено, что начало развития новой части обусловливается присутствием на поверхности разреза одного из тех органов, которые принимают участие в образовании данной части. Если у головастика из поверхности среза удалена из хвоста спинная струна, то новый хвостуле регенерирует (рис. 102 с). Если, однако, конец спинной струны подходит ближе к месту разреза, то путем образования новых клеток, которые впоследствии {184} достигают обнаженной поверхности, может начать развиваться хвост (рис. 102 а—f).
Если у дождевого червя из разреза удалена брюшная нервная цепочка, то новая голова на передней поверхности разреза не развивается, а может образоваться несколько кзади, в том месте, где заканчивается старый нервный ствол (рис. 103). Регенерация ноги саламандры из отрезанного конца зависит от присутствия какого-то вещества, происходящего из кости. Если вместо того, чтобы отрезать ногу, расщепить ее с одной стороны до кости, то этим может быть вызвано развитие другой вторичной ноги. Эти особые органы — нервный ствол, спинная струна и периост — могут быть названы организаторами новой части, так как они должны присутствовать для того, чтобы могло регенерировать новое целое. Не так давно это же явление было обнаружено в некоторых опытах по прививке эмбриональных организаторов. Это послужит темой следующей главы.
| << | {185} | >> |
Самым замечательным фактом в эмбриональном развитии является правильная последовательность изменений, протекающих от стадии яйца до взрослого организма. Это особенно ясно видно в стадиях дробления яиц аннелид и моллюсков, где каждая клетка может быть прослежена вплоть до определенного органа зародыша. Эта связь настолько характерна, что дробление считалось протекающим как бы по заранее намеченному пути, т. е. целесообразно. Однако такое чисто телеологическое утверждение с научной точки зрения является нелепостью.
Может показаться, что теория локализации органообразующих участков во время дробления есть современная форма очень старой доктрины о преформации. Эта древняя доктрина просто постулировала наличие в яйце (или в спермин) очень маленького человечка, которому надо только вырасти большим, чтобы стать человеком. Мы в настоящее время уже слишком много знаем о том, что происходит при развитии яйца, для того чтобы принять такое простое объяснение. Современное учение о преформации или прелокализации в яйце органообразующих участков тоже ничего не говорит нам о том, каким образом каждый участок яйца обособляется для того, чтобы стать определенной частью зародыша, даже если таковая связь строго закономерна. Имеются экспериментальные данные, показывающие, что это представление не принимает во внимание важные движения веществ в яйце и последующие влияния, которые тоже так же существенны для процессов развития, как и более простая связь региональной детерминации. Кроме того, взгляд о прелокализованных участках не учитывает потенций яйца до определения его медиальной плоскости, и очень может быть, что подобные же возможности имеются и на более поздних стадиях развития. Игнорирование {186} этих фактов может привести к упущению некоторых из наиболее существенных особенностей процесса развития. Образование целого эмбриона из части яйца или из изолированного бластомера показывает, что допущение строгой прелокализации органообразующего участка иногда не объясняет происходящих явлений.
Последние открытия показали, что некоторые важные фазы развития начинаются влиянием одного участка на другой. Эти факты не позволяют нам согласиться с положением, что вопрос был бы разрешен, если мы смогли бы проследить орган зародыша до определенного участка оплодотворенного яйца. Сейчас мы должны рассмотреть результаты экспериментальных исследований, имевших прямое отношение к этим вопросам.
Существует два близких по своему характеру рода опытов, которые дают важные факты в этой области, — это, во-первых, искусственный перенос клеток из одного участка эмбриона на другой участок того же самого или другого зародыша для того, чтобы выяснить, самодиференцируются ли там эти клетки или же они принимают участие в развитии той части, на которую они были перенесены. Во-вторых, перенос некоторых веществ в другие участки, где они вызывают развитие органов, которые в противном случае в этих участках не появились бы. Большинство из этих опытов было проведено на относительно крупных яйцах хвостатых амфибий, некоторые же из них — на мелких яйцах морских ежей.
Если небольшая часть эктодермы тритона, состоящая из поверхностных клеток, взята на стадии ранней гаструлы с участка впереди бластопора, т. е. из той области, которая нормально стала бы частью нейральной пластинки и трансплантирована на другую гаструлу в участок, где нормально развилась бы эктодерма (рис. 104 d—f), то она становится эктодермой. Обратно, когда кусочек из эктодермальной области трансплантируется (рис. 104 а—с) на другую гаструлу в область проспективной нейральной пластинки, то он принимает участие в образовании нейральной пластинки. Очевидно, что в это время судьбу трансплантированной части определяет ее локализация, из чего может следовать, что эктодерма в это время либо еще не диференцирована, а если и диференцирована, то не изменена настолько сильно, что не может быть обратима другими влияниями, когда находится в новом положении.
| {187} |
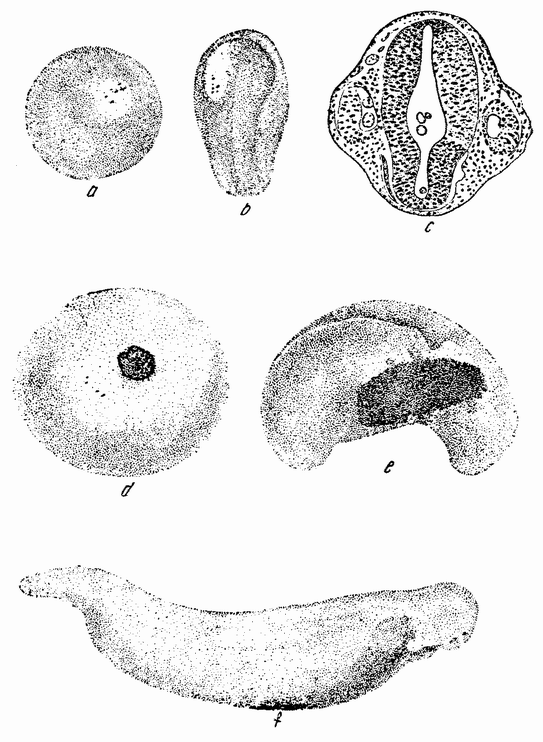 |
|
Рис. 104. а — ранняя гаструла Triton taeniatus, в которую в ооласть проспективной нейральной пластинки пересажен маленький кусочек проспективной эктодермы Т. cristatus; b — более поздняя стадия а; с — поперечный разрез через зародыш, который развился из двух предыдущих стадий. Из имплантированного кусочка развилась правая сторона нейральной трубки и правый глазной пузырек; d — ранняя гаструла Т. cristatus, в которую в область проспективной эктодермы был пересажен кусочек вещества проспективной нейральной пластинки Т. taeniatus. На более поздних стадиях (е и f имплантант становится частью поверхностной эктодермы (по Шпеману). |
| {188} |
С другой стороны, если подобное изменение происходит на немного более поздней стадии, когда впереди бластопора обрисовывается нейральная пластинка, то кусочек нейральной пластинки, перенесенной на вентральную поверхность другого зародыша, погружается внутрь и образует пузырек, стенки которого диференцируются в нейральную трубку. Обратно, кусочек вентральной эктодермы, перенесенной на нейральную пластинку, не становится частью нейральной трубки. В последнем случае можно сказать, что диференциация зашла настолько далеко, что клетки не передиференцировываются и не принимают региональных черт своего нового местоположения. Эти опыты, хотя и не выясняют природу этих влияний, но они тем не менее показывают, что изменения прогрессивны и до некоторого момента могут быть обратимы. Даже если, как это постулирует теория мозаичности, в яйце и была предетерминация, то из опытов очевидно, что вначале более важен эффект положения.
Не менее важны и другие опыты, определяющие направление развития некоторых частей. Первый опыт этого рода показывает, что хрусталик глаза лягушки развивается в соответствии с наличием под эктодермой глазных пузырьков (выростов мозга). Это было показано двумя путями: во-первых, отрезанием конца глазного пузырька и имплантацией его под эктодерму соседнего участка на голове. Эктодерма вокруг имплантанта при этом утолщается и образует хрусталик; во-вторых, эктодерма участка, на котором должен был бы появиться хрусталик, удалялась и на обнаженный участок помещался новый кусочек эктодермы с другой части тела. Из этого кусочка тоже развивался хрусталик. Очевидно, хрусталик развивается в зависимости от наличия под ним глазного пузырька. Можно сказать поэтому, что хрусталик индуцируется в эктодерме присутствием оптической трубки: последняя является для хрусталика индуктором или, как выражаются в последнее время, организатором.
Аналогичные данные были получены при изучении развития центральной нервной системы тритона. Для того чтобы понять значение этих фактов, можно напомнить прелокализацию частей эмбриона у тритона, изображенную на рис. 105 А—Е. На рисунке А зародыш изображен сбоку, а на рис. В показан его вентральный вид сейчас же после образования дорзальной губы. Материал, {189} образующий нейральную пластинку, занимает верхний дорзальный квадрант, а будущая хордальная энтодерма и мезодерма лежат в квадранте под нейральной пластинкой. Последние участки ввернутся внутрь, обходя дорзальную губу бластопора. Судьба этих клеточных слоев была обнаружена путем маркирования краской различных мест на поверхности зародыша, благодаря чему име-
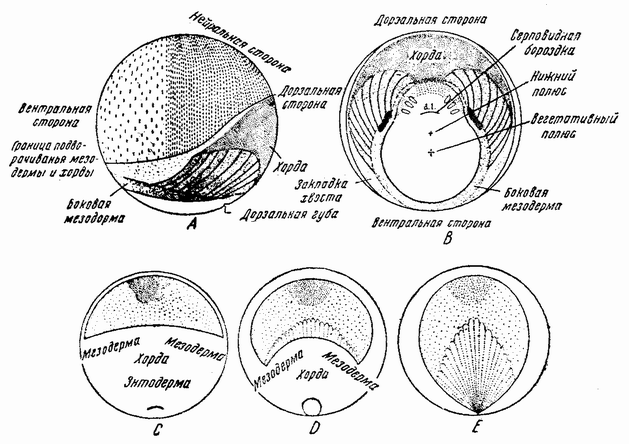 |
|
Рис. 105. А и В — схемы участков поверхности зародыша тритона сейчас же после появления дорзальной губы гаструлы; А — вид сбоку; В — вид с желточного полюса (по Фогту). На трех нижних рисунках С, D и Е изображен участок проспективной нейральной пластинки (пунктир), хорды и мезодермы впереди дорзальной губы. На этих рисунках видно также удлинение материала нейральной пластинки во время закрывания бластопора, в то время когда хорда и мезодерма вворачиваются в губу бластопора (по Гертлеру). |
лась возможность проследить передвижения этих участков по мере последующего развития.
Самыми важными опытами были те, в которых вещество непосредственно спереди бластопора (рис. 106 а, проспективная хордальная энтодерма) и с боков его (проспективная мезодерма) было удалено и имплантировано на вентральную или латеральную поверхность другого, {190} более молодого зародыша, т. е. находящегося еще на стадии поздней бластулы или в начале гаструляции. Имплантант погружается при этом под поверхность, а затем вокруг и перед ним в эктодерме хозяина развивается неиральная складка (рис. 106 b). Могут даже появиться органы зрения, слуха и другие родственные структуры. Сам имплантант дает начало спинной струне и мезобластным сомитам. Про-
 |
|
Рис. 106. а — схема гаструлы тритона. Был удален кусочек поверхностной энтодермы нейральной пластинки этой нейрулы, а извлеченный кусочек дорзальной стенки первичного кишечника был введен в бластоцель молодой гаструлы. Эмбрион, развившийся из этой гаструлы, и его трансплантант изображены на рисунке b. Небольшая вторичная неиральная трубка находится сбоку переднего конца первичной нейральной трубки (по Гейнитцу). |
спективная хордальная мезодерма действует как индуктор, вызывая развитие в эктодерме нейральной трубки, которая в свою очередь может индуцировать в индиферентной эктодерме появление органов слуха, носовых ямок и других структур, т. е. органы, которые при нормальном развитии не появились бы в этих участках. Являются ли проспективная энтодерма, спинная струна или мезодермальные клетки стадии гаструлы равнопотенциальными, не вполне ясно из одного этого опыта, так как, как это изображено на рисунках, {191} между этими клетками на стадии гаструлы нет такой резкой разграничительной линии. Эти первые опыты дали основание предполагать, что индукция идет от клеток хордальной энтодермы, которая нормально вворачивается внутрь и становится крышей первичного кишечника, над которым у нормального эмбриона развивается нейральная пластинка. Наличие такого же рода реакции следовало бы предполагать и в опыте, в котором хордальная энтодерма оказывается лежащей где-либо под эктодермой. Однако этот простой и, казалось бы, логический вывод оказался слишком ограниченным для того, чтобы охватить все, что известно в этой области, ибо другие опыты показали, что индукция не является свойством одной хордальной энтодермы, а им обладают также другие клетки молодого зародыша.
Следующие опыты были проделаны другим способом, а именно путем вставки кусочков из проспективной нейральной пластинки (взятых на различных стадиях) в бластоцель других эмбрионов, находящихся на стадии ранней гаструлы (рис. 107 и 108). Группа клеток в области проспективной нейральной пластинки была удалена с крыши ранней стадии гаструлы (в то время когда бластопор имеет U-образную форму) и помещена внутрь сегментационной полости молодой гаструлы (рис. 107 а, b). По мере протекания гаструляции введенный кусочек оказывается лежащим напротив крыши или краевой зоны реципиента, так как масса желточных клеток вдавливается внутрь и заполняет первичную сегментационную полость. Кусочек лежит, следовательно, между эктодермой снаружи и желтком или мезодермой изнутри. В общем можно сказать, что эктодермальный трансплантант сливается с поверхностной эктодермой хозяина и не действует как организатор, но в некоторых случаях индуцирует нейральную трубку. Из этого следует, что на этой стадии эти эктодермальные клетки, т. е. клетки проспективной нейральной пластинки, не всегда вызывают изменения в хозяине, как это делают подобные клетки, взятые из более поздней стадии.
В другой серии опытов из проспективного переднего конца поздней гаструлы был удален кусочек эктодермы с желточной пробкой (рис. 107 с) и перенесен в сегментационную полость более молодой гаструлы (рис. 107 d). Результаты были изменчивыми; иногда над прививкой появлялась только выпуклость, но в других случаях имелись {192} явные следы индукции, как это видно из развития структур нейральной пластинки (8 случаев из 29).
В третьей серии опытов был взят кусочек из переднего угла нейральной пластинки (рис. 108 a) и введен в сегментационную полость более молодой гаструлы (рис. 108 b); в этом случае над ним в «вентральной» эктодерме хозяина развивается нейральная пластинка. Кроме того, на перед-
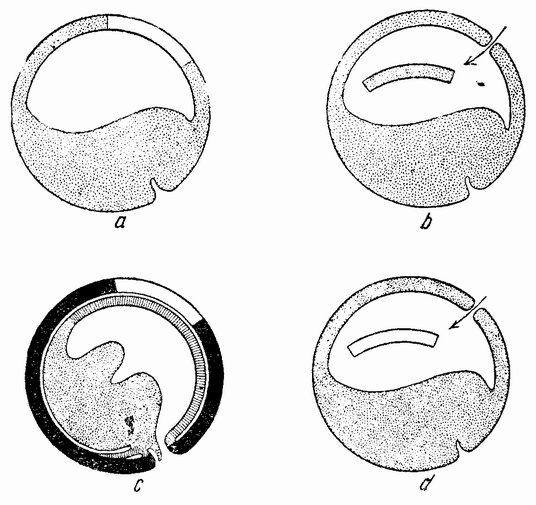 |
|
Рис. 107. Схемы, иллюстрирующие трансплантацию частей гаструлы тритона, а — кусочек верхней стенки пересаженный в b; с — кусочек внешнего слоя верхней поверхности, трансплантированный в d (по Мангольду). |
нем конце нервной трубки могут развиться вторичные глазные бокалы, а из лежащей сверху эктодермы — хрусталик; могут также появиться слуховые пузыри и носовые ямки, которые иногда развиваются на переднем конце вторичного эмбриона.
В предыдущем опыте трансплантированный кусочек происходил из передней части нейральной пластинки. {193} Главными появлявшимися при этом органами были органы, характерные для переднего конца. Однако если частица взята из заднего конца нейральной пластинки более взрослого эмбриона, то эти передние органы отсутствуют и из тканей хозяина развивается хвостовая область. Имеются дальнейшие данные, что ответ хозяина в некоторой степени зависит от того, где расположен пересаженный кусочек, — ближе к передней области, дальше назад или в стороны. На результат влияет также стадия хозяина, на которой ему был привит кусочек. Другими словами, не только все части проспективной эктодермы хозяина
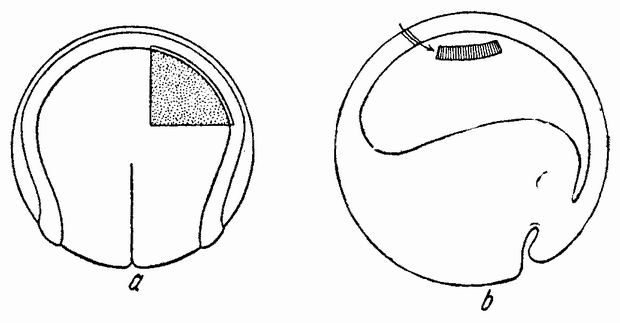 |
|
Рис. 108. Кусочек эктодермы из правого переднего конца нейралыгой пластинки зародыша тритона (а), трансплантированный в сегментационную полость молодой гаструлы (b) (по Мангольду). |
не одинаково отвечают на один и тот же индуктор, но и наоборот, влияние самих индукторов зависит от стадии зародыша или участка, из которого они происходят.
Новейшие опыты с организаторами у амфибий далеко отбросили виталистическое содержание термина организатор, которое могло бы вкладываться в него при выборе этого слова. Эти опыты показывают, что индукция может быть непосредственно обусловлена химической или физической реакцией между прививкой и хозяином, ибо эти эксперименты показали, что и мертвый трансплантант может вызвать индукцию, т. е. действовать как «организатор» или индуктор.
Если кусочки [проспективной эктодермы, нейральной пластинки или даже кусочки неоплодотворенного яйца убиваются нагреванием (до 60 °) и вводятся в бластоцель {194} молодой гаструлы, то они индуцируют развитие нейральной пластинки в проспективной эктодерме гаструлы. Путем вторичной индукции могут также развиться органы зрения, слуха и равновесия и т. д. Результаты индукции оказы-
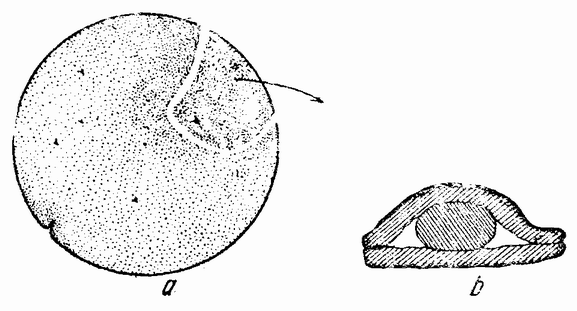 |
|
Рис. 109. Кусочек верхней стенки молодой гаетрулы (а), помещенный между двумя лепестками живой эктодермы (b) (по Гольфтретеру). |
ваются совершенно такими же, как и при употреблении живых индукторов.
Такого же рода результат, хотя и не такой сильный, может быть получен другим путем. Если кусочек из молодой нейральной пластинки или дорзальной губы бластопора убивается нагреванием в солевом растворе и затем
 |
|
Рис. 110. Кусочек поверхностного слоя молодой гаструлы из а, помещенный на мертвое вещество из другого яйца (b) по (Гольфтретеру). |
окружается двумя листками проспективной эктодермы, как это показано на рис. 109, то в последней развивается нейральная трубка, а позднее и характерная нейральная структура. Для протекания этого процесса существенно наличие индуктора, ибо если в солевой раствор помещаются {195} одни листки эктодермы, то они образуют только эктодерму. Индукция может быть вызвана также и другим образом. Частицы нейральной пластинки, предположительной эктодермы (рис. 110 а) или даже энтодермальные клетки убиваются нагреванием в солевом растворе. На них помещаются кусочки эктодермы (рис. 110 b), которые при соприкосновении с субстратом дольше чем один день развивают нейральные трубки (рис. 111 а—b). Эти резуль-
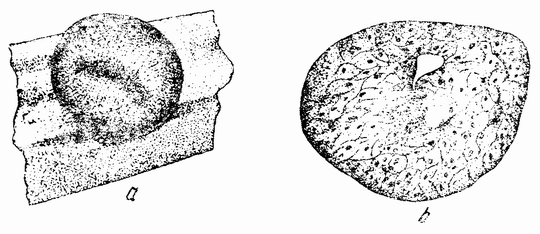 |
|
Рис. 111. а — образование ненральной трубки в кусочке, лежащем на убитом веществе, как на рис. 110; b — разрез через то же самое (по Гольфтретеру). |
таты показывают, что способность индуцировать нейральную трубку имеется и в мертвом веществе. Простейшее объяснение этого заключается в том, что индукция вызывается химической реакцией.
Обратная реакция, а именно влияние хозяина на прививку, лучше обнаруживается в других опытах, особенно же при переносе молодых слуховых пузырьков и молодых зачатков конечностей в другие области зародыша.
Первые опыты касались слуховых пузырьков. Они появляются по одному с каждой стороны головы сначала в виде пластинки или чаши клеток из внутреннего слоя эктодермы, которая по мере ее погружения в лежащую под ней соединительную ткань закругляется в полый пузырек, из которого развиваются три полукружных канала, вестибулум, лабиринт и другие части внутреннего уха (рис. 112). На более поздних стадиях ухо является уже системой с тремя измерениями и имеет наружно-внутреннюю, дорзо-вентральную и передне-заднюю оси. {196}
Как только появился слуховой пузырек, покрывающая его эктодерма может быть приподнята и пузырек удален. Он может затем быть перемещен на то же место, но с повернутыми тем или иным образом осями, или транспланти-
 |
|
Рис. 112. а — молодой зародыш лягушки; виден лоскуток эктодермы, приподнятый для того, чтобы обнажить ушной пузырек под ним; b — разрез через образующийся ушной пузырек; с — трансплантированный ушной пузырек, повернутый дорзо-вентрально; d — нормальный правый пузырек; е — полностью развитое нормальное внутреннее ухо (по Шпеману). |
рован на то же место, но с другой стороны эмбриона, повернутым или не повернутым, или же помещен где-либо в теле зародыша. Результат этих опытов несколько {197} затемнен, благодаря тому, что пузырек перед или во время его диференцировки поворачивается. Но в случаях, когда такого вращения не произошло, слуховой пузырек оказывается самодиференцирующимся, т. е. он развивается по своим собственным осям и не зависит от своего отношения к хозяину.
Сходные проблемы были изучены на зачатках конечностей амфибий, которые могут быть удалены и трансплан
 |
|
Рис. 113. Схема, показывающая ориентировку привитых зачатков конечностей (по Гаррисону). |
тированы в том же положении с той же самой стороны зародыша или с противоположной с одной или двумя измененными осями, либо же перенесены на другие участки зародыша. Как показано на рис. 113, эти зачатки появляются в виде местных утолщений во внешнем слое мезодермы, которая образует сердцевину зачатка. Покрывающая эктодерма может быть приподнята и зачаток удален. Он может быть затем помещен под эктодерму в другом участке или с противоположной стороны другого эмбриона на место удаленного с этой стороны зачатка. Возможные {198} ориентировки для переднего и заднего зачатков показаны на рис. 113 (внизу). Кружком изображен на рисунке зачаток, а буквами внутри кружков его оси. Буквы вне кружков обозначают оси хозяина. Опуская детали, можно сказать, что в момент этих опытов зачаток конечности был «поляризован» только в одном передне-заднем направлении. Это значит, что в каком бы новом положении зачаток ни находился, его развитие находится в соответствии с этой господствующей осью, тогда как развитие в отношении двух других осей определяется его новым положением,
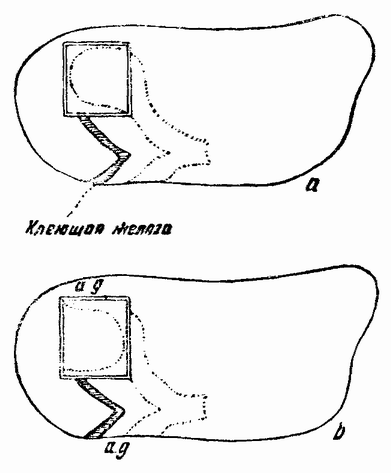 |
|
Рис. 114. Схема, показывающая местоположение удаленного кусочка (а), повернутого на 180° и перемещенного, как показано на рис. 6. Клейкая железа служит для ориентировки. Пунктирная линия на обоих рисунках показывает очертания будущей жаберной крышки (по Экману). |
т. е. влиянием, исходящим от хозяина. Присутствуют ли во время пересадки и эти две последние поляризации и изменяются ли они последующими влияниями хозяина или же они еще не развились в зачатке — на этот вопрос этот опыт ответа не дает. Однако, повидимому, у некоторых видов осевые отношения могут быть фиксированными на более ранней стадии по сравнению с другими видами. Вероятно также, что у определенных видов дорзо-вентральная и латеральная оси тоже настолько детерминированы {199} на более поздних стадиях, что в новом положении их реверсия не будет иметь места.
Опыты с жаберными щелями лягушек показали, что впячивание эктодермы, которое встречается и сливается
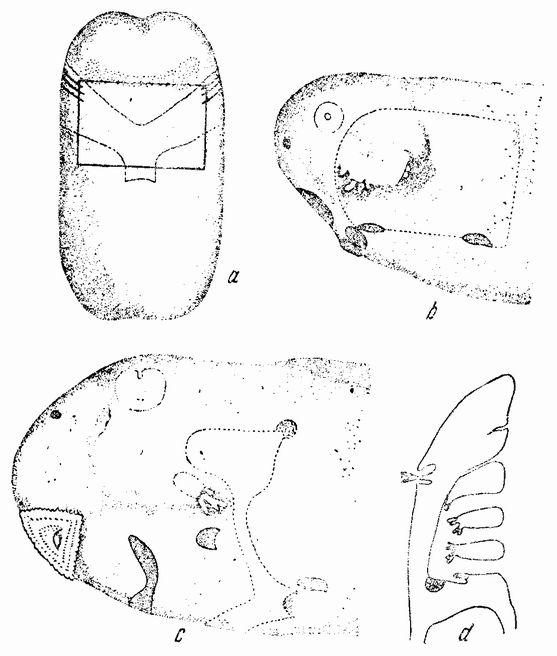 |
|
Рис. 115. а — вентральный кусочек эктодермы, трансплантированный на обнаженный жаберный участок b. Как видно на рнс. d, в трансплантанте индуцированы жаберные щели; жаберный участок покрыт крышкой (с и d) (по Экману). |
с выпячиванием энтодермы глотки, индуцируется складкой энтодермы. Так, например, когда у молодого эмбриона удален квадратный кусочек эктодермы (рис. 114), который лежит в проспективной жаберной области, и пересажен, {200} будучи повернутым на 180°, на сходный участок другого зародыша, у которого предварительно был удален квадратный участок эктодермы такой же величины, то в прививке в эктодермальном выпячивании позднее появляются жаберные щели. Этот опыт не вполне убедителен, потому что кусочек повернут на 180°, что приводит вертикальное впячивание, хотя и в повернутом виде, но все же приблизительно, в то же самое положение, что и раньше. Кроме того, многое в опыте указывает на то что ко времени операции некоторая индукция уже имела место.
Более удовлетворительные результаты были получены при удалении кусочка эктодермы с вентральной поверхности над сердцем, как показано на рис. 115 а, и при пересадке его на обнаженную жаберную область другого эмбриона (рис. 115 b). Жаберные щели возникают в эктодерме прививки над лежащими под ними энтодермальными мешками. Позже на дугах появляются жабры (рис. 115 d). В придачу в трансплантированном кусочке появляется несколько мелких жабр на переднем краю прививки (рис. 115 с), которые могли быть индуцированными перед прививкой, так как боковые края вентрального кусочка эктодермы могут накрывать жаберную область. Но даже и в этом случае могла иметь место некоторая индукция,исходящая от хозяина, так как жабры не появлялись на заднем краю того же кусочка, лежащего позади жаберной области.
Еще более убедительны опыты удаления прямоугольного кусочка, простирающегося позади жаберной области в область головной почки, поворачивания его на 180° и имплантирования на тот же обнаженный участок зародыша, как показано на рис. 116. В этом случае область прививки, из которой образовались бы жаберные щели, лежит теперь позади жаберной области, как это показано вертикальными пунктирными линиями на рис. 116 a, тогда как эктодерма прививки, лежащая над будущими энтодермальными мешками, нормально лежит позади этого участка. Эктодермальные впячивания появляются позднее над энтодермальными мешками жаберной области и на них образуются жабры, как показано на горизонтальном разрезе на рис. 116 с. В этом случае имеется ясное доказательство индукции.
Будет ли эктодерма с любой части зародыша одинаково хорошо реагировать в таких опытах — сомнительно. В одном случае, в котором эктодерма из задней области была {201} помещена над жаберным участком, не развились ни щели, ни жабры. Здесь, как и в некоторых других опытах, имеются указания о наличии для каждого органа градиента в передне-заднем, дорзо-вентральном и, может быть, в радиальном направлении, который простирается кнаружи от проспективного центра, понижаясь с увеличением расстояния. Это, вероятно, может быть объяснено тем, что изменения в направлении будущего развития каждого участка про-
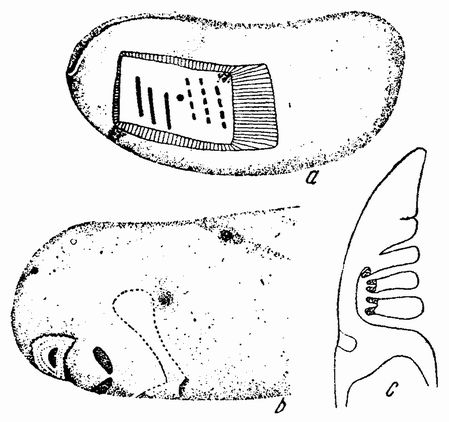 |
|
Рис. 116. а — кусочек поверхностной экзодермы над жаберным участком, достигающий уровня головной почки, удален и повернут в передне-заднем направлении; жаберные щели индуцируются в том конце прививки, который является теперь передним; прерывистые линии обозначают место, в котором должны были бы развиться жаберные щели; b — крышка, покрывающая жаберный район; с — на разрезе показаны индуцированные жаберные щели (по Экману). |
исходят, начиная с очень ранней стадии. Если эти изменения не зашли слишком далеко, то они могут исчезнуть или быть измененными в обратном направлении, если участок привит в новые условия окружающей среды. Сущность этих проспективных влияний в настоящее время совершенно неизвестна и в различных частях эмбриона она может быть весьма различной как по своему характеру, так и по количественному выражению. {202}
Различные комбинации половинок двух яиц морского ежа Paracentrotus могут быть получены следующим образом. После удаления яиц из их оболочки им дают развиваться до 16-клеточной стадии. Наличие микромеров на вегетативном полюсе дает затем возможность определить плоскость разделения групп бластомеров. Для того чтобы сделать это, яйца за десять минут до разделения, которое производится с помощью стеклянной иглы, помещаются в морскую воду, не содержащую кальция.
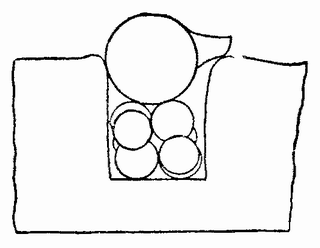 |
|
Рис. 117. Схема, показывающая, как получается сращивание вместе бластомеров морского ежа (по Герстадиусу). |
Когда полярность обеих соединяемых половинок одинакова, как показано на рис. 118 a, из них развивается нормальный плутеус. Так как разделение могло пройти либо через проспективную среднюю плоскость, если таковая в это время уже определена, либо под прямым углом к этой средней плоскости, то, повидимому, в это время такая плоскость отсутствует или, если присутствует, то может быть изменена и установлена заново. Когда половинки соединяются так, как показано на рис. 118 b, т. е. с обратной полярностью, то вегетативная область каждой половинки инвагинируется, как изображено на рис. 119 а и b. В этом случае образуется плутеус с двумя совершенно разделенными пищеварительными трактами, с двумя лежащими друг против друга анальными отверстиями, двумя средними и передними кишками и с одним общим ротовым отверстием (рис. 119 с). Скелет почти удвоен. {203}
Если половинки соединены, как изображено на рис. 118 с, т. е. ориентированы под прямым углом друг к другу, то гаструляция начинается в двух точках и два первичных кишечника раньше или позже соединяются, образуя раздвоенную или единую кишку. В этом случае развивается либо типичный, либо, более часто, ненормальный плутеус.
 |
|
Рис. 118. Схема различных способов соединения половинок 16-клеточных стадий морского ежа (по Герстадиусу). |
При соединении анимальных половинок, показанном на рис. 118 d, развивается бластула с двумя рядами ресничек, идущими в противоположных направлениях. Если соединены (рис. 118 е) меридиональная и анимальная половинки, то комбинация состоит из 12 мезомеров (эктодерма), 2 макромеров и 2 микромеров и развивается типичный плутеус (рис. 120 с). {204}
Если анимальная половинка присоединена к анимальному же концу целой 16-клеточной стадии (рис. 118 f), то такая комбинация содержит две трети эктодермы и одну
 |
|
Рис. 119. а — гаструляция соединенных половинок (рис. 118); b — то же самое у другого подобного же соединения; с — развившийся двойной плутеус (по Герстадиусу). |
 |
|
Рис. 120. а — атипичный плутеус, образовавшийся в результате соединения, изображенного на рис. 121 с; b — плутеус, развившийся из соединения изображенных на рис. 121 d; с — плутеус из комбинации е рис. 121 (по Герстадиусу). |
треть энтодермы, из нее может развиться плутеус, размеры которого будут крупнее нормального.
В случае соединения вегетативной половинки с анимальным концом целой 16-клеточной стадии (как показано на рис. 118 d) получается один плутеус с двумя кишечниками и одним ртом, сходный с изображенным на рис. 119 с. {205}
При соединении двух вегетативных половинок (рис. 118 h) происходит две инвагинации, причем личинки оказываются при этом совершенно ненормальными.
Четыре микромера имплантировались в другую 16-клеточную стадию (рис. 118 i), образованную из двух половинок, как изображено на рис. 118 а. Число мезенхимных клеток на стадии гасгрулы было удвоенным. Развивались типичные личинки без удвоения скелетных элементов.
В предыдущих случаях внутренние поверхности соединенных половинок были направлены друг к другу.
 |
|
Рис. 121. Схема различных способов соединения половинок 16-клеточных стадий морских ежей (по Герстадиусу). |
В другой серии опытов наружная поверхность одной половинки обращалась к внутренней поверхности другой половинки (рис. 121 а). Для того чтобы понять, как эта система выправляется, полезны будут следующие схемы. На рис. 122 а нарисована нормальная бластула, у которой вегетативный полюс заштрихован. Если эту половинку отрезать и повернуть так, чтобы ее поверхность расположилась напротив внутренней поверхности верхней половинки (рис. 122 b), то обе половинки изгибаются, как изображено на рис. 122 с. Затем две полости соединяются путем удаления перегородки (122 d) и получается единая полость (е—f). Внешние поверхности обеих {206} половинок находятся теперь на внешней поверхности одной бластулы, но здесь имеется два центра вегетативных клеток, лежащих на экваторе друг против друга.
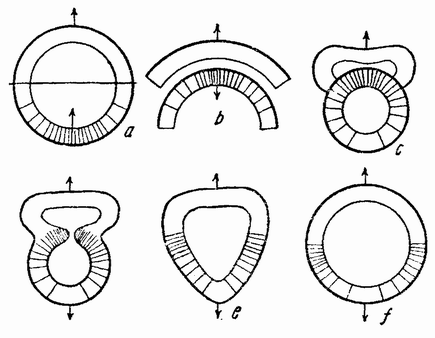 |
|
Рис. 122. Схемы, показывающие результат соединения вегетативных половинок, повернутых и соединенных с анимальной половинкой так, как показано на рис. b (по Герстадиусу). |
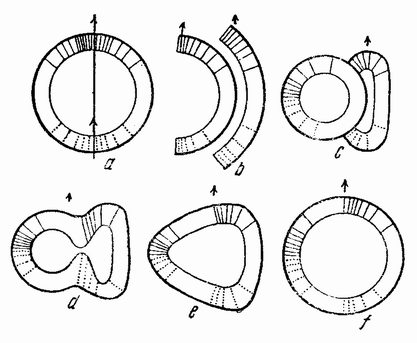 |
|
Рис. 123. Схема, показывающая результат соединения меридиональных половинок 16-клеточных стадий, как показано на рис. b (по Герстадиусу). |
На рис. 123 а представлены две разделенные по анимальной оси половинки, одна из которых перевернута и {207} ее внешняя поверхность помещена внутрь другой половинки (123 b). Обе половинки закругляются, как показано на рис. 123 с. Перегородка удаляется (d) и получается единая полая бластула. Вегетативная область находится на той же стороне бластулы, но несколько в сторону от нее (d—f).
С помощью этих схем легко понять поведение соединенных половинок 16-клеточной стадии. Если две половинки соединены, как на рис. 121 а, получаются как типичные, так и атипичные плутеусы. Если две половинки соединены, как на рис. 121 с, то развиваются тоже и типичные и атипичные плутеусы. При обратной комбинации (рис. 121 d) развитие идет совершенно неправильно (рис. 120 b). Некоторая регуляция в этих случаях происходит, но гаструляция нарушается и плутеусы оказываются ненормальными в отношении энтодермы (рис. 120 b).
Если восемь клеток двух вегетативных полушарий соединяются с той же ориентировкой (рис. 121 b), то происходит две инвагинации, а получающиеся плутеусы оказываются очень ненормальными.
Если с каждой из двух половинок удаляются микромеры и половинки соединяются затем так, как изображено на рис. 121 f, то не образуется первичной мезенхимы, а с опозданием появляется вторичная мезенхима. В этом случае может развиться нормальный плутеус.
Эти опыты прививок показывают, что верхняя и нижняя половинки уже специфичны в отношении их будущего развития. Этого можно было ожидать из поведения анимальной и вегетативной половинок при изолировании их, а также и из поведения анимальных и вегетативных половинок несегментированных яиц. Существует некоторое взаимное восстановление клеток, но в целом при наличии двух групп макромеров очевидна тенденция удвоения (что дает начало первичному кишечнику). В какой степени макромеры действуют в качестве индуктора для остального эмбриона, не вполне ясно. Факты говорят скорее противоположное.
| << | {208} | >> |
В определении пола, так же как и в очень многих других биологических явлениях, часто может казаться, что один и тот же конечный результат достигается различными путями. У одних групп животных имеется хромосомальный механизм, приводящий к образованию равных количеств самцов и самок. У других же групп развитие семенников или яичников определяется не различиями в хромосомальном строении, а внешними и внутренними факторами другого порядка. В обоих случаях локализация половых клеток, будь то яичники или семенники, в определенных областях тела является, повидимому, строго эмбриологической проблемой, имеющей то же основание, что и локализация других органов. Итак, возникает вопрос, является ли именно локализация органа — семенника или яичника — первоначальным фактором, определяющим пол, или же главную проблему составляет развитие семенных или яйцевых клеток. У гермафродитных форм, обладающих и семенниками, и яичниками в различных частях одной и той же особи,вопрос обстоит, повидимому, так же, как и для всех эмбриологических локализаций, и это, повидимому, правильно также для тех раздельнополых животных, у которых семенник и яичник расположены в теле несколько различно.
Эта проблема более трудна для анализа в тех случаях, когда семенники и яичники занимают у самца и самки практически одно и то же положение. Последующее обсуждение будет касаться определения пола главным образом с точки зрения проблемы локализации, а не самого механизма определения пола. Однако сначала следует кратко описать последний. {209}
У некоторых групп позвоночных, у большинства насекомых, а также у некоторых круглых червей, морских ежей и др. известно существование хромосомного механизма, регулирующего число самцов и самок. Этот механизм состоит из особых хромосом, называемых половыми хромосомами. Самка имеет две половых хромосомы (XX), самец же одну Х-хромосому и обычно другую хромосому, являющуюся партнером первой и называемую Y-хромосомой. Эта хромосома отличается от всех других хромосом тем, что в ней либо полностью, либо частично отсутствуют факторы, связанные с определением пола. Ее иногда назы-
 |
|
Рис. 124. Образование гамет у самки типа XX и самца ХУ. Один сорт яиц и два сорта сперматозоидов. |
вают пустой, или индиферентной.Точнее,Y-хромосома либо вовсе не содержит генов, подобных содержащимся в других хромосомах, либо по крайней мере их в ней не так много. Самец XY продуцирует два сорта сперматозоидов с Х-хромосомой и с Y-хромосомой. Яйцо, содержащее Х-хромосому, оплодотворенное Х-спермием, дает самку (XX); Х-содержащее яйцо, оплодотворенное Y-спермием, дает самца (XY). Этот процесс все время повторяется (рис. 124).
Существует также и обратный механизм определения пола, при котором имеется два сорта зрелых яиц и только один сорт спермиев (рис. 125). Самка имеет строение WZ, а самец — ZZ. Здесь Z и W обозначают половые хромосомы. Z — яйцо, оплодотворенное Z-спермием, дает самца; W-яйцо и Z-спермий дают самку. Пустой {210} является здесь W-хромосома. Этот механизм обнаружен у бабочек, птиц и некоторых рыб.
Общие во всех этих случаях черты заключаются в том, что гены пола лежат в хромосомах, причем не только в половых, но также и в других хромосомах, называемых собирательно аутосомами. Факты показывают, что здесь замешано много генов, лежащих как в половых хромосомах, так и в аутосомах, причем определяющим пол фактором является баланс между этими генами. Таким образом, можно сказать, что при типе XX—XY Х-хромосома содержит больше женских генов, а аутосомы — больше мужских. Y-хромосома же является индифе-
 |
|
Рис. 125. Образование гамет у самки типа WZ и самца ZZ. Два сорта яиц и один сорт сперматозоидов. |
рентной. Эта теория не является лишь предположением, основанным на присутствии двух Х-хромосом у самок и одной у самца, а покоится на фактах нарушения нормальных отношений в результате случайного присутствия необычного хромосомного комплекса, как это наблюдается иногда у триплоидов, интерсексов, суперсексов и т. п.
При типе WZ—ZZ отношение между половыми хромосомами и аутосомами предполагается принципиально таким же, как и при типе XX—XY, но в этом случае принимается, что половые хромосомы (ZZ) содержат больше мужских генов, а аутосомы — больше женских.
Теория баланса между генами, участвующими в определении пола, принципиально не отличается от применяемой к наследственности вообще, когда каждый признак рассматривается как продукт многих генов, {211} присутствующих во всех или во многих хромосомах. Изменение в любом из этих генов создает новое равновесие, приводящее к иному конечному продукту. Пусть, например, изменение в единственном гене приводит к образованию нового признака или группы признаков. Это не означает, что признак вызывается самим измененным (т. е. мутировавшим) геном, а лишь то, что, вследствие установления нового равновесия, этот ген действует только как диференциатор в процессах, образующих некоторые признаки. Ударение здесь делается именно на диференциальном эффекте одного гена или пары их. Может показаться вероятным, что различия между самцом и самкой тоже обусловливаются единственным диференцирующим геном, но, как было указано выше, факты говорят другое. Это положение не означает, что все гены, участвующие в определении пола или в образовании любых других признаков, одинаково эффективны. Даже один из этих генов может в некоторых случаях изменить соотношение, но в определении пола это равновесие является, повидимому, конечным результатом деятельности многих генов. И действительно, пол связан с различиями не в единственном органе, а с целым комплексом различий, многие из которых взаимно связаны и взаимно зависимы.
Можно сказать, что во всех тех случаях, где имеется особый механизм определения пола, пол особи определяется в момент оплодотворения, несмотря на то, что истинное развитие половых отличий может и не проявляться до более поздних стадий. Это не значит, что даже в этих случаях пол не может быть изменен или даже обратим в той или другой своей особенности, а означает лишь, что при обычных для каждого вида условиях жизни этот механизм дает равное количество самцов и самок. При особых же внешних и внутренних условиях процессы развития могут быть нарушены и результат изменен. Как бы ни были интересны эти исключительные случаи, все же наличие их не влияет на интерпретацию того, что происходит в нормальной внешней среде раздельнополых видов, у которых существует хромосомный механизм определения пола.
Иногда говорят, что самка потенциально является самцом, а самец есть потенциальная самка. Это говорит нам, однако, не больше, чем факты сами по себе, так как особи, имеющие все сорта генов, оказываются сходными во {212} всем за исключением их генного баланса. Подобные утверждения делаются иногда с очень глубокомысленным видом и предназначаются для того, чтобы создать впечатление, что «пол» является физиологическим явлением, которое должно изучаться как таковое независимо от генетической ситуации. Если цель такого рода утверждений бросить тень на хромосомный механизм определения пола, то они не достигают цели, так как в настоящее время имеются многочисленные факты, показывающие, что такой механизм в нормальных условиях производит равные числа самцов и самок. Но каким образом генный баланс производит этот результат — это является в такой же мере физиологической проблемой, как и путь, при помощи которого внешняя среда может физиологически влиять на определение пола особи.
Поскольку имеется различие между генетическим определением пола и определением пола внешней средой, последний метод имеет преимущество в том отношении, что он может дать возможность подойти к вопросу не чисто аналитически, а более непосредственно со строго количественными методами. Недавнее открытие мужских и женских гормонов у позвоночных и получение этих веществ в чистом виде могут приблизить нас к генетической теории. Но идентификация этих гормонов с генами пола является сильно преувеличенной. Гормоны, как и все другие признаки, несомненно являются косвенным продуктом генов, но они действуют такими окольными путями, что могут быть скорее отнесены к факторам внешней среды, чем к факторам, непосредственно действующим на гены той клетки, в которой они образуются. Некоторые из этих веществ, влияющих на половые протоки или на вторично половые признаки, вырабатываются в семенниках или яичниках (если не в самых половых клетках), но их действие обычно сказывается на других органах тела и в таких случаях может становиться частью проблемы развития этих органов.
Половые клетки — яйца и спермин — развиваются в строго локализованных областях эмбриона. Их происхождение может быть в некоторых случаях прослежено до первых стадий дробления, а в отдельных случаях до единственной клетки, но, как было уже сказано выше, локализация {213} органа, в котором они находятся, не представляется отличной от проблемы локализации всех других органов. Но происходит ли независимое определение характера развивающихся в органе половых клеток равновесием генов — является открытым вопросом. Может казаться, что у гермафродитного животного, у которого в одном участке тела развивается семенник, а в другом яичник, характер половых клеток всецело определяется тем же самым фактором, который определяет положение органа. Но имеются также и такие гермафродитные организмы, например, подобные устрице, у которых одна и та же гонада является попеременно то семенником, то яичником. Различие между этими состояниями представляет для эмбриолога особенно интересную проблему и требует дальнейшего анализа.
У такого гермафродитного животного, как дождевой червь, у которого семенники возникают в одной части тела, а яичники в другой, повидимому, те же самые факторы, которые локализуют семенники, определяют и развитие в них семенных клеток, причем эта же зависимость имеет место и для яичников. У форм, подобных дождевому червю, повидимому, нет хромосомальных различий, как у однополых животных, но тем не менее это дает право предполагать, что между генами, вызывающими развитие спермиев и яиц, существует равновесие. Можно думать, что этот баланс такого рода, что в одних органах он изменяется одним образом, а в других — иначе. Этот аргумент может быть применим также и для однополых животных, у которых имеется хромосомальный механизм определения пола. У некоторых из таких организмов семенник и яичник оказываются расположенными в одной и той же части тела, но разовьется ли орган в яичник или семенник, может определяться генным балансом. Последнее утверждение может быть выражено и иначе. Местоположение половых органов может быть одинаковым у обоих полов, но орган может в каждом случае состоять из двух частей. Генное равновесие может тогда определять, которая из этих частей разовьется первой — яичник у WZ-особи или семенник у ZZ-особи. Однажды развившись, каждая часть может вырабатывать гормон, который подавляет другую часть. Это объяснение может быть иллюстрировано развитием половых органов у птиц. У птиц, как указывалось выше, имеется хромосомальный {214} механизм определения пола (WZ—ZZ). У самки становится функционирующим, т. е. вырабатывает яйца, только один яичник, лежащий с левой стороны тела. Правый же «яичник» остается в рудиментарном состоянии. Если же у молодой птицы удален левый яичник, то правый яичник развивается в семенник и из основания удаленного левого яичника тоже может развиться семенник. Оба семенника могут вырабатывать сперму. Практически не подлежит сомнению, что клетки, дающие начало регенерирующим семенникам, у таких овариотомированных птиц имели ту же хромосомную формулу, что и яйца. Несмотря на их хромосомную идентичность, примордиальные клетки в условиях нормального развития самки развиваются в яйца, а после овариотомии того же организма — в спермин. Более того, место расположения обоих органов оказывается приблизительно одинаковым. Как же мы можем объяснить этот кажущийся парадокс?
Никаких затруднений для формального объяснения этого парадокса нет. Если эмбрион цыпленка в начале развития имеет хромосомальное строение самки (WZ), то его хромосомный баланс на молодых стадиях определяет развитие яичников. При наличии же хромосомального строения самца развиваются семенники. Можно сказать, что орган, который в соответствии с формулой пола развивается первым, своим развитием подавляет развитие другого органа. С удалением же яичника это подавляющее действие исчезает. Почему же тогда предпочтительно развиваются не семенники, а новый яичник? Ответ, который может быть дан на этот вопрос, заключается в том, что в новых условиях внешней среды равновесие оказывается более благоприятным для развития семенников. Правда, такое объяснение представляет собой только немного больше, чем констатацию фактов, но тем не менее оно все-же центрирует внимание на самом важном моменте, а именно, что новая реакция зависит от внешних условий, в которых она производит свой эффект. Вначале среда молодой птицы дает преимущество для развития яичника; среда более старой птицы благоприятствует началу развития латентного семенника. В несколько более разработанном виде этот аргумент был бы таким: во время первой закладки органов размножения в будущей самке генное равновесие ведет к более быстрому развитию той части первичного органа, которая станет яичником. Яичник подавляет {215} всякое дальнейшее развитие тестикулярной части первичного органа. Наоборот, в молодом эмбрионе самца генный баланс оказывается таким, что создается преимущество для развития семенника, который подавляет развитие овариальной части первичной гонады. Когда яичник самки удаляется, его подавляющее действие на рудимент семенника исчезает и, несмотря на генетическое строение клеток, тестикулярная часть начинает теперь продолжать свое развитие, потому что ей, так сказать, был дан уже толчок в этом направлении.
Кастрирование самца птицы не вызывает изменения в органе, который может регенерировать. Если какая-либо регенерация и происходит, то развиваются новые семенники. Обычно если кастрация полная, то семенник не всегда развивается, потому что рудимент его тоже удален. Когда же кастрация не полная, то семенник развивается из оставшегося куска. Можно предполагать поэтому, что при оперировании самца удаляется первичный рудимент органа, если таковой имеется, или же если у самца вначале присутствовал рудимент яичника, то он может быть полностью подавлен развитием семенника.
Теоретически регенерировавший семенник овариотомированной самки должен продуцировать два сорта зрелых половых клеток, подобно тому, как это получается с яйцами, имеющими сходное хромосомальное строение, а именно часть спермиев должна иметь одну Z-хромосому, а часть — W-хромосому. У птиц этого не было обнаружено, но имеются данные, что у других форм, у которых существуют подобные же отношения, хромосомальный механизм соответствует ожидаемому.
Известно несколько случаев, в которых пол особи непосредственно определяется средой.
Гефирея Bonnelia является самым известным примером влияния среды на определение пола. Самка этого вида имеет тело величиной примерно со сливу с хоботом, длина которого около метра. Самец же представляет собой маленькое червеобразное существо в несколько миллиметров длиной, которое живет, как паразит, в матке самки. Оплодотворенные яйца после откладки развиваются в свободно плавающую трохофорную личинку (рис. 126, a). Если какие-нибудь из этих личинок сядут на хобот взрослой самки, {216} то они становятся самцами (рис. 126, с). Личинки, оставшиеся свободно плавающими, опускаются, в конце концов, на дно и постепенно превращаются в самок (рис. 126, b). В этом случае среда определяет не только развитие яичника или семенника, но и развитие других органов мужских и женских особей тоже в различной степени зависит от внешних условий. Самцы во многих отношениях организованы более просто и остаются более близкими к личиночной форме. Так как они в этом же состоянии достигают половозрелости, то их можно отнести к неотеническим формам. Данных о наличии у Bonnelia хромосомального механизма определения пола нет, а если он отсутствует, то нет оснований предполагать, что у нее происходит превращение пола в том или другом направлении. Факты скорее говорят за то, что в одних условиях среды индиферентная личинка развивается непосредственно в самца, а в другой среде в самку. Опыты показали, что на хобот взрослой самки садятся не предетерминированные самцы, а что любая личинка может в этих условиях стать самцом. Правда, среди сотен личинок, содержимых в морской воде, некоторые могут стать самцами, но только спустя некоторое время. Такой же результат может обусловливаться и создавшимися в культурах искусственными условиями, например, повышением кислотности или щелочности; возможно также,что самцами могут стать голодавшие личинки. В таких культурах часть личинок становится также интерсексуальными, т. е. особями, обнаруживающими некоторые вторичнополовые признаки самца и самки (рис. 126, d—f). Интерсексы могут быть также получены путем перенесения личинок в морскую воду, после того как они пробыли некоторое время на хоботе самки.
Нормально личинки остаются прикрепленными к хоботу самки в течение приблизительно ста часов, после чего они заползают в переднюю кишку самки и через две или три недели становятся половозрелыми самцами. В первые полтора дня пребывания на хоботе передний конец личинки укорачивается, теряет свой пигмент и глазные пятна; на второй день быстро размножаются половые клетки, которые станут впоследстви испермиями, на третий день появляется выводной проток и его воронка; сперматозоиды развиваются на шестой день. Если личинки удалены с хобота, то степень этих изменений зависит от продолжительности {217} пребывания на нем личинок. Опыты показали, что пребывание на хоботе от семи до десяти дней достаточно, чтобы образовалось мужское состояние. Когда личинки через короткое время удаляются с хобота в воду, то полу-
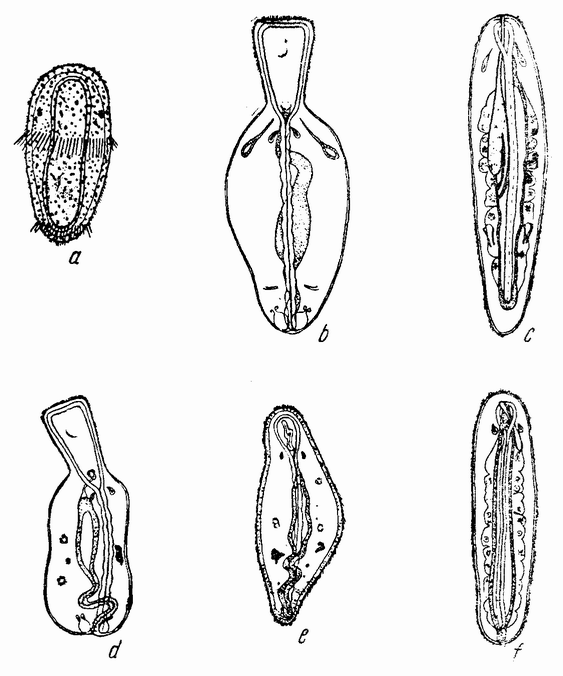 |
|
Рис. 126. Зародыши Bonneha, а — индиферентная личинка; b — женская личинка; с — мужская личинка; d — женская интерсексуальная личинка, е — промежуточная личинка; f — мужской интерсекс (по Бальцеру). |
чаются следующие результаты: личинки, остававшиеся на хоботе только от двадцати до тридцати часов и затем удаленные, становятся только слегка подобными самцам в тех органах, которые реагируют первыми; те личинки, {218} которые оставались на хоботе только десять часов, менее похожи на самцов; остававшиеся только четыре часа вообще почти не обнаруживают мужских признаков. В общем можно сказать, что после короткого пребывания на хоботе изменение после удаления имеет место только в переднем конце и пропорционально продолжительности пребывания личинки на хоботе. Это соотношение показывает, что некоторые влияния требуют большего времени, чем другие, или, скажем иначе, реагирование различно для разных органов. Затронутый здесь вопрос заключается не в простом противопоставлении мужского состояния женскому, а касается каждого органа в отдельности, так как одни органы отвечают на воздействие более четко, чем другие. Эта проблема была бы слишком упрощена, если ее рассматривать только как изменение в половом состоянии, ибо, кроме половых клеток, оказываются затронутыми многие органы и притом независимо от половой железы. В нормальных условиях молодое животное остается на хоботе самки достаточно долго для того, чтобы изменить все свои органы в органы, характерные для самца.
Опыты показывают, что влияние хобота является химическим. Если куски хобота или кишечника высушить и затем вымочить в морской воде, то полученный раствор будет превращать индиферентных личинок в самцов или по крайней мере изменять их в мужском направлении. При небольшом подкислении морской воды (0,5 см3 n/10 HCl + 20 см3 морской воды) развитие индиферентных личинок тоже идет в мужском направлении (91,6%). Имеется, следовательно, достаточно оснований для вывода, что здесь участвуют химические агенты, при этом любопытно, что секрет хобота имеет щелочную реакцию. Кроме того, если эта секреция слишком сильна, то она даже убивает личинки. Можно, хотя для этого и нет еще достаточных оснований, заключить, что как кислота, так и секреция действуют как подавители, не допускающие, так сказать, развития женских органов.
Существуют некоторые предположения относительно путей, с помощью которых постулированные мужские и женские гены могут участвовать в этих результатах. Но до сих пор эти предположения не вышли за пределы чисто умозрительных стадий. Данные, полученные при изучении Bonrielia, не требуют, повидимому, для своего объяснения гипотез, которые были бы основаны на точке зрения, что {219} здесь происходит такое определение пола, при котором самцы и самки являются результатом действия механизма половых хромосом. Если, как это кажется вероятным, у Bonnelia нет механизма половых хромосом, определяющего равновесие генов после оплодотворения, то совершенно бесполезно пытаться объяснить эти результаты предположением, что индиферентные личинки являются уже либо самцами, либо самками.
В других случаях, в которых было обнаружено существование механизма половых хромосом, имеются данные, по-
 |
|
Рис. 127. Личинка Perla marginata слева). Яйце-семенник молодого самца (справа) (по Юнкеру). |
казывающие, что этот механизм может быть неэффективным на ранних стадиях развития одного пола, но на более поздних стадиях может вести к развитию мужских или женских органов. Например, было показано, что у мужской личинки Perla marginata одновременно присутствуют типичные яичник и семенник (рис. 127), тогда как у женской личинки развивается только яичник. Самец в этом случае имеет строение XY, а самка — XX. Эти факты могут быть истолкованы таким образом, что равновесие хромосом у самок таково, что оно непосредственно {220} вызывает у них развитие яичника; на молодых же стадиях самца это равновесие недействительно, благодаря чему развиваются и семенник, и яичник. Но на более поздних стадиях развития самца равновесие генов или, быть может, присутствие семенника приводит к подавлению яичника.
Другая иллюстрация найдена у мухи Miastor (рис. 128), у которой определение пола происходит по формуле XX—XY. Оплодотворенные яйца этой мухи превращаются в личинок, которые, оставаясь еще личинками, продуцируют яйца; у личинок образуются только яйца и они развиваются партеногенетически. Может иметь место длинный ряд последовательных поколений партеногенетических
 |
|
Рис. 128. Miastor, самец и самка (справа). Три личинки с молодью внутри (слева) (по Кале). |
личинок. Изменение в среде может в любое время заставить личинок продуцировать яйца, которые дают начало крылатым самцам и самкам. Яйца этих мух, для того чтобы развиться, должны быть оплодотворены спермой самца. Было обнаружено существование двух линий личинок: одна из них дает начало только взрослым самцам, другая же — взрослым самкам. Из этого следует, что партеногенетическими яйцами воспроизводятся как генетически мужские, так и генетически женские личинки. Очевидно, на личиночных стадиях равновесие между хромосомальными факторами не мешает самцам XY продуцировать на этих стадиях яйца, но, когда среда изменяется, развитие самца или самки определяется равновесием хромосом. Из обширной литературы, касающейся определения пола у лягушек и жаб, здесь может быть приведено только несколько типичных примеров. В Европе имеются две расы {221} травяной лягушки (Rana temporaria), у одной из которых головастики развиваются непосредственно в самцов или в самок. Эта раса называется диференцированной расой. У другой расы все головастики вначале кажутся «самками» или по крайней мере они содержат в гонадах крупные яйцеобразные клетки. У этой расы у половины головастиков развиваются впоследствии семенники, замещающие собой первые стадии развития яиц, тогда как у другой половины головастиков яйца продолжают развиваться. Нельзя быть вполне уверенным в том, что у самца этой лягушки имеется XY-пара хромосом, хотя факты определенно говорят в пользу этого состояния. Если это так, то можно сказать, что хромосомальный механизм определения пола на ранних стадиях будущего самца как таковой не функционирует, так как сначала появляются яйца; однако на более поздних стадиях равновесие между Х- и Y-хромосомами прямо или косвенно определяет продуцирование сперматозоидов. Должна ли переходная стадия самцов быть названной интерсексуальной, является предметом определения.
В связи с приведенными выше рассуждениями некоторый интерес представляет собой эмбриональное развитие гонад. На дорзальной стенке полости тела очень рано появляются два генитальных валика, которые в поверхностном слое содержат первичные половые клетки. Каждый валик состоит из наружного коркового и внутреннего медулярного вещества. На более поздних стадиях в корковом веществе у самок образуются яйца, а в медулярном веществе у самцов — семенные клетки. И те и другие происходят из первичных половых клеток. У видов, самцы которых имеют не прямое развитие, яйца появляются сначала в корковом веществе, а затем, когда в медулярном веществе развиваются семенные клетки, они исчезают. Поэтому создается впечатление, что судьба первичных половых клеток или их преемников определяется той частью органов размножения, в которой они лежат.
Интересные результаты, имеющие значение для этих отношений, были получены в опытах, в которых молодые головастики, до того как развились их половые органы, были привиты друг к другу своими боковыми сторонами. Такие, так называемые парабиотичесие, близнецы могут быть либо парой самцов, либо парой самок, либо самцом и самкой. Так как соединение было сделано прежде, чем {222} можно было различать пол, то на основании случайного комбинирования ожидается такое соотношение комбинаций: одна пара самцов, две пары состоящих из самца и самки и одна пара самок. При изучении этих пар на более поздней стадии оказалось, что они обнаруживали соединение именно в этой пропорции. Был также обнаружен интересный факт: оказалось, что в некоторых случаях, в которых были соединены разнополые головастики, развитие половых органов могло быть нарушено. У жаб никакие органы не затрагиваются этим влиянием, но у лягушек такая независимость оказывается верной только для ранних стадий, впоследствии же «конфликт» между яичником и семенником становится очевидным и чем теснее близнецы соединены, тем сильнее выражен этот эффект. Сначала воздействие начинает обнаруживаться на внутренних членах каждой пары гонад, которые лежат более близко друг к другу, чем члены наружной пары. Семенник господствует над развитием яичника в том отношении, что он имеет тенденцию подавлять корковый слой яичника. Подавление корковой части ведет к развитию медулярной части яичника, т. е. той части, которая дает начало семенным клеткам. Однако в исключительных случаях положение оказывается обратным и в таких случаях предполагается, что яичник либо уже вначале опередил семенник, либо же он развивается скорее, чем семенник. Здесь замедленным оказывается развитие медулярной части мужской железы одного из близнецов Эти результаты создают впечатление, что в развитии обеих частей семенника молодой личинки имеется конкуренция — развитие коркового вещества подавляет развитие медулярной части органа, и наоборот. Какая часть начинает развиваться первой у нормального головастика, окончательно определяется генетическим строением особи: если это самец (XY), то преимущество имеет медулярный слой (за исключением начала развития); если самка, то — корковый слой. Но эти нормальные отношения могут быть нарушены условиями среды, которые могут вызвать частичное превращение пола. Только что описанные опыты прививок иллюстрируют это. Подобный же результат может быть вызван и температурным воздействием. Например: если молодые головастики древесной лягушки содержатся при температуре в 20°, то развитие семенников и яичников идет обычным путем и оба пола возникают в отношении 1:1. Но если молодые головастики перенесены в более высокую {223} температуру (32°), то семенники продолжают развиваться нормально, а яичники оказываются затронутыми — их корковое вещество останавливается в развитии и яйца дегенерируют. Медулярное вещество начинает тогда быстро расти. Развиваются характерные половые канатики семенников и, наконец, появляются семенные клетки самца. Следовательно, подавление наружного слоя позволяет развиться внутренней части половой железы, что ведет к образованию семенников. Здесь мы приближаемся к полному знанию отношений между развитием мужской и женской половых желез. В большинстве случаев та или другая железа в зависимости от генетического строения особи начинает развиваться первой, но этот элемент времени может быть обнаружен условиями среды. Если сначала развивается корковое вещество — получается яичник, если медулярное вещество — возникает семенник. Развитие каждого из них подавляет развитие другого.
Экспериментальные данные, полученные из другого источника, доказывают, что у лягушки самка имеет строение XX, а самец — XY. Изредка находятся взрослые особи, у которых часть одной и той же гонады является семенником, а другая часть — яичником. Эти лягушки являются генетическими самками. В одном случае оказалось возможным оплодотворить яйца диференцированной расы травяной лягушки спермой такого гермафродита. Все потомки от этого скрещивания оказались женскими. Если гермафродитная самка имеет строение XX, т. е. является генетической самкой, то каждый зрелый сперматозоид должен содержать одну Х-хромосому. Каждое яйцо использованной в этом опыте самки должно тоже содержать по одной Х-хромосоме; поэтому ожидается, что все потомки должны иметь строение XX, т. е. будут самками. Яйца гермафродита тоже были оплодотворены спермой самца диференцированной расы. От этого скрещивания получились самцы и самки в равных количествах, что опять-таки находится в соответствии с XX—XY формулой. Когда яйца гермафродита были оплодотворены спермиями от той же особи, то почти все потомство, за исключением нескольких интерсексов, было женским.
У рогатого скота давно было известно любопытное явление, которое теперь может быть объяснено на основании результатов, полученных в опытах с парабиотическими близнецами. Когда рождаются телята-близнецы, из которых {224} один нормальный самец, а другой самка, то последняя обычно стерильна. Такого теленка называют фримартином. Наружные гениталии фримартина — женские или по крайней мере более похожи на женские, чем на мужские, но репродуктивные органы напоминают семенники. Было обнаружено, что каждый из этих близнецов происходит из отдельного яйца и что позднее кровеносная система обоих эмбрионов сообщается посредством анастомозов, образующихся между эмбриональными оболочками (рис. 129). Факты убедительно показывают, что фримартин начинает развиваться как генетическая самка и что связь с мужским эмбрионом посредством циркуляции крови по-
 |
|
Рис. 129. Два зародыша коровы с соединенными плацентами; один из этих зародышей станет фримартином (по Лилли). |
давляет развитие яичника, в результате чего развиваются тестикулярные структуры. Происходит частичное превращение пола.
Вопросы «интерсексуальности» интенсивно изучались у шелковичной бабочки. В различных частях света — в Европе, Азии и Японии — существуют различные расы этой бабочки. Внутри каждой из этих рас число самцов и самок оказывается равным. Принимается, что у шелковичной бабочки, так же как и у других бабочек, существует механизм половых хромосом, причем самка имеет строение WZ, а самец ZZ. Если самка европейской расы скрещивается с: японским самцом, то получается равное число самцов и самок, но при противоположном скрещивании сыновья оказываются нормальными, а дочери интерсексами. Эти интерсексы представляют собой мозаики из мужских и женских частей. Различные расы японской бабочки при {225} скрещивании с европейскими расами или с какой-либо другой дают различные комбинации нормальных полов и интерсексов. В одной серии скрещиваний все до одной самки были изменены в самцов. Они были женскими интерсексами. В других же скрещиваниях все самцы были изменены в самок. Это были мужские интерсексы. Эти результаты могут быть объяснимы, исходя из гипотезы, что различные расы имеют различное половое равновесие. Существуют сильные женские расы, слабые расы и т. д. Имеется некоторая нея:ность относительно того, где это равновесие существует — между половыми хромосомами и аутосомами, между W- и Z- хромосомами или же между цитоплазмой и всей группой хромосом.
Мозаичный характер интерсексов объясняется Гольдшмидтом с помощью гипотезы, что результат определяется общим количеством мужского и женского энзимов, присутствующих в эмбрионе во время закладки мужских или женских частей. Так, например, при некоторых комбинациях половых факторов этих расовых гибридов вначале присутствует больше мужского энзима и поэтому особь начинает свое развитие, как самец, но на более поздних стадиях превалирующим становится женский энзим, количество которого обгоняет выработку мужского энзима, и поэтому последующие органы оказываются похожими на таковые у самок. При других комбинациях получается обратный результат. Это в некотором смысле является повторением уже наблюдавшихся фактов, которые интерпретировались в духе половых энзимов, развивающихся в клетках затронутой части, а не в самих половых железах, как это имеет место у позвонодных.
Эта гипотеза ставит ребром вопрос о том, действуют ли все гены в течение всего времени или же одни из них активны на одной стадии, а другие на другой. Если мы предположим, что все гены активны с самого начала развития, то генное равновесие и получающееся в результате его отношение энзимов в нормальном женском эмбрионе (WZ) будет таково, что все органы, закладывающиеся в течение развития, оказываются женскими; в мужском же эмбрионе (ZZ) все органы являются с самого начала развития мужскими. Но у гибридов шелковичной бабочки получается новое генное равновесие, которое по мере протекания развития вызывает изменение в пропорции энзимов и в половых признаках. Итак, становятся возможными два или {226} даже три альтернативных объяснения (1). Согласно взгляду, что все гены действуют все время, должно быть предположено, что у нормальных рас при наличии определенного генного равновесия все органы, отличающиеся у обоих полов, затронуты в мужском направлении, а при наличии другого равновесия, те же самые органы затронуты в женском направлении. Но у гибридной комбинации органы, закладывающиеся первыми, реагируют в одном направлении, а закладывающиеся позже — в противоположном направлении к тому же самому равновесию. Здесь подчеркивается характер реагирования разных органов на различных стадиях развития, причем надо отметить, что реагируют вещества в самих затронутых клетках. Предполагается, что генное равновесие остается все время тем же и изменяющийся результат зависит от специфического pear ирования соматических или даже половых органов к чему-то, что вырабатывается в клетках (2). При предположении, что некоторые гены, вследствие ли своего числа или свойств, более активны на одной стадии развития, а другие гены — на другой, должно быть принято, что у гибридов тот или иной набор генов оказывается более активным на ранних стадиях, т. е. предоминирует, а гены другого набора становятся более активными на более поздних стадиях. Пользуясь энзимологическими терминами можно сказать, что на вырабатывающееся количество самого энзима влияет стадия развития эмбриона (3). Имеется еще и третья возможность, которая является своего рода комбинацией первых двух, а именно, что результат зависит непосредственно от скорости развития самих этих энзимов в различных случаях. В некоторых гибридных комбинациях мужской энзим вырабатывается сначала в избытке, вследствие чего и закладывающиеся сначала органы оказываются мужскими; впоследствии же женские энзимы перекрывают мужские и развитие поворачивается в женском направлении. Результат, следовательно, зависит от скорости или быстроты, с которой вырабатываются эти два энзима. Следовательно, все гены деятельны все время, но у различных комбинаций гибридов различно время, потребное для того, чтобы мужской или женский энзим оказался в избытке. Здесь ударение сделано на скорости развития энзима независимо от эмбриональных стадий. При первой же точке зрения (1) ударение делалось на реагировании рано и поздно закладывающихся органов с {227} общим количеством энзима, выработанного генами; при второй точке зрения (2) изменение зависит от времени, в течение которого контрастирующий набор генов становится более активным, и, наконец, при третьей точке зрения (3) — от скорости, с которой тот или иной энзим вырабатывается, причем эта скорость определяется не различными генами, а скоростью развития энзимов в эмбрионе. В настоящее время мы не можем остановиться на какой-либо одной из этих возможностей. Любая из них может представляться как физиологический процесс, но так много факторов еще не известно, что претендовать на какую-нибудь из них как на динамическую теорию пола кажется слишком смелым; ибо динамика, в физическом смысле, этого термина имеет дело с измеримыми количествами и известными физическими свойствами рассматриваемых агентов.
| << | {228} | >> |
В предыдущих главах почти все внимание было обращено на происходящие в течение развития необратимые изменения формы, потому что эти изменения лучше всего видны и являются наиболее изученными признаками развития. Наряду с этими изменениями формы происходит образование различных специфических веществ, причем как изменения формы, так и химические изменения можно рассматривать как физиологические процессы. Взаимоотношение тех и других изменений ставит нас лицом к лицу с некоторыми фундаментальными вопросами. Как это обычно понимается, физиология имеет дело с периодическим функционированием органов, которое происходит без какого-либо постоянного изменения в их форме или структуре. Возникает вопрос, являются ли происходящие в эмбрионе изменения чем-либо большим или чем-либо отличным от физиологических функций взрослого организма.
Однако рискованно считать за доказанное, что взрослый организм как таковой структурно находится в статическом состоянии. Известно, что многие органы взрослого животного непрерывно обновляются; кожа снашивается с поверхности и ее клетки замещаются новыми клетками из более глубоких слоев; в железах тоже происходит, повидимому, замещение функционировавших в течение определенного времени клеток, клетки крови непрерывно разрушаются и обновляются. Наиболее постоянными клетками, сохраняющимися в течение всей жизни, являются центральная нервная система, мышцы и кости, но иногда и кость может обновляться, и после поранения ее периоста или наружного слоя она обнаруживает замечательную способность регенерации новой костной структуры. Тем не менее различие остается: новый орган сохраняет свою форму, а форма эмбриона претерпевает быстрые изменения. {229}
В других же отношениях имеется очевидное сходство. Дыхание имеет место как у эмбриона, так и у взрослого организма, причем в обоих случаях кислород поглощается, а углекислота освобождается; вода играет важную роль и тут и там; обоими используются углеводы для некоторых химических изменений. То же самое касается жиров и протеинов. Взрослый организм получает все эти вещества извне и переваривает и метаболизирует их, но в яйце они уже присутствуют, и трансформация протеинов, жиров и углеводов, вероятно, происходит путем одинаковых химических изменений как в яйце, так и у взрослого организма. Превращение этих веществ обусловлено присутствием энзимов, которые, действуя как катализаторы, перерабатывают вещество, снабжая эмбрион энергией, необходимой для его превращений. Запасные вещества яйца используются как пища для быстрого роста эмбриона и они, следовательно, превращаются в протоплазму. Для развития важны также многие соли. Для эмбрионов некоторых морских животных, развитие которых протекает в морской воде, было показано, что многие из этих солей они находят непосредственно в море. В яйцах, развивающихся в воде как в пресной, так и в соленой, содержится также вода. У наземных животных, особенно у насекомых, в яйцах тоже содержится достаточно воды для того, чтобы поддержать эмбрион в течение первых стадий развития. Потеря воды, вследствие испарения, предотвращается окружающей яйцо оболочкой. Наконец, практически всегда в яйце присутствуют витамины. Последних хотя и немного, но все же достаточно для поддержания первых стадий развития. Может ли яйцо или эмбрион вырабатывать их из своих собственных веществ, как это делают многие растения, — проблематично, но можно с уверенностью предположить, что эмбрион содержит их в достаточном количестве, чтобы поддержать свое развитие до тех пор, пока эти вещества не смогут получаться им извне. На более поздних стадиях необходимые витамины воспринимаются несомненно с пищей, как у взрослых животных.
Имеется, следовательно, достаточно данных, показывающих, что существует химия развития, сравнимая во многих отношениях с химией жизнедеятельности взрослого организма. Остается еще вопрос, вправе ли мы предполагать, что познание этой химии будет достаточным для {230} объяснения претерпеваемых эмбрионом изменений формы. Было бы одинаково рискованно на основании тех или иных соображений как утверждать, так и отрицать это. Физические свойства яйца и эмбриона также играют роль в развитии. Более того, яйцо само по себе имеет определенную, хотя и лабильную, собственную организацию, служащую основой для его развития.
Заслуживает внимания тот факт, что видимые созидательные процессы развития могут быть сведены к многократно повторяющимся небольшим изменениям. Деление яйца имеет все признаки клеточного деления клеток тканей взрослых животных и многих одноклеточных форм. Растворение ядерной стенки и распад хроматиновой сетки на составляющие ее хромосомы, митотические фигуры в цитоплазме и т. п. происходят при делениях как яйцевых, так и не яйцевых клеток. В одном отношении деление яйца отличается от деления тканевых клеток — это скоростью последовательных делений, но даже здесь сходство велико, так как между двумя последующими делениями имеется покоящееся ядро. Только одни бактерии при наличии обильной пищевой среды по скорости своего деления приближаются к скорости деления яйца. Не может быть сомнений, что яйцо потому делится быстро, что в нем самом заключается запас необходимых веществ в готовой для утилизации форме. В другом отношении дробление яйца отличается от дробления клеток тела. При дроблении яйца не происходит увеличения размеров дочерних клеток до исходных, прежде чем наступит следующее деление. Это также может быть с уверенностью приписано обилию веществ, стимулирующих деление и дающих необходимую энергию. Это положение относится особенно к первым делениям яйца и менее верно для более поздних стадий.
Прилегание друг к другу клеток дробящегося яйца после каждого деления может быть имитировано с помощью прижатых друг к другу с периферии капелек масла. Несомненно, в обоих случаях играет роль поверхностное натяжение, но если от капелек масла удалена надавливающая поверхность, то они расплываются в стороны. При удалении же с яйца оболочки оплодотворения бластомеры остаются попрежнему вместе. Надо думать поэтому, что что-то, кроме поверхностного натяжения, удерживает их; в оболочке есть нечто, склеивающее их вместе и возможно притягивающее подобные клетки друг к другу. Было показано, что в воде, {231} не содержащей кальция, бластомеры не склеиваются и могут быть отделены друг от друга. Это означает, что присутствие кальция в морской воде необходимо для удержания этих клеток вместе.
Амебоидное движение отдельных клеток, характерное для некоторых процессов развития, напоминает движение белых кровяных телец взрослых животных и по всем признакам подобно движению свободно живущих амеб. Было сделано много предположений относительно физической стороны амебоидного движения, и хотя нельзя претендовать на то, что оно вполне понятно, все же вероятно по крайней мере, что такие движения связаны с изменением поверхностного натяжения.
Известно много примеров втягиваний зародышевых листков эмбриона. Процессы инвагинации или гаструляции являются прекрасными примерами этого. Крупные клетки одной полусферы полости бластулы по достижении определенной стадии вворачиваются внутрь (рис. 33 а, b, с). Если изучить отдельные вворачивающиеся клетки, то окажется, что каждая имеет широкий внутренний конец и узкий внешний. Это могло бы случиться, если бы они были вдавлены внутрь внешней силой, но так как такие сллы неизвестны, то, повидимому, в это время происходит изменение формы отдельных клеток. Расширение их внутренних концов напоминает распластывание выдвигающегося конца амебы, указывая этим, что поверхностное натяжение здесь тоже играет роль. Это может быть объяснено предположением, что увеличение углекислоты или каких-либо других веществ, понижая поверхностное натяжение внутренних краев клеток, вызывает их расширение. Поэтому вворачивание стенок происходит как чисто физический процесс. Правда, для этого требуется, чтобы клетка была везде гомогенной, что, вероятно, не так. Могут быть и несомненно существуют различия между внутренним и наружным концами клеток, которые берут свое начало из структуры яйца, но даже если это так, то стимул, действующий на внутренний конец, может быть все-таки химическим агентом.
Примеров инвагинаций и выпячиваний очень много. Мешки возникают в некоторых случаях из стенки первичного кишечника; глазные стебельки выпячиваются из переднего отдела мозга; глазные бокалы и слуховые пузыри представляют собой выпячивания по бокам головы {232} и т. д. Все они возникают, претерпевая изменения одного и того же рода.
В стенках некоторых первоначальных органов могут возникать местные утолщения. Они вызываются локальным увеличением числа клеток, которое сопровождается передвижением отдельных клеток с поверхности. Процесс этот весьма сходен с впячиванием слоя, однако здесь клетки действуют индивидуально, а не как слой, и этот процесс является, повидимому, лишь видоизменением впячивания или, наоборот, последний процесс есть видоизменение первого.
Погружение нейтральной пластинки для образования трубки — центральной нервной системы позвоночных — другой пример продольного впячивания вместо радиального по отношению к центру, как это происходит при гаструляции. Утолщение клеток частично обусловлено вталкиванием клеток из поверхностного слоя, что сопровождается одновременным закручиванием внутрь или инвагинацией всей пластинки как целого. Движения всей пластинки, по-видимому, совершаются таким же путем, как и в других случаях впячивания.
Открытие того, что при наличии под поверхностью организатора, даже когда такой организатор состоит из мертвого вещества, инвагинация центральной нервной системы может совершаться не в тех областях, где она происходит обычно, много дает для доказательства того, что этот процесс есть физическая реакция, происходящая в результате химического стимула.
Предыдущие случаи представляют собой прекрасные примеры наиболее характерных процессов эмбрионального развития, связанных с изменением формы. В них, повидимому, нет ничего таинственного, т. е. ничего такого, что не может быть связанным с физическими принципами, которые не кажутся недосягаемыми для экспериментального анализа. Эмбриолог будет, хотя бы временно, удовлетворен, когда он сможет точно показать пригодность физического объяснения для таких изменений формы.
Но это только часть истории развития и картина эта, вероятно, слишком проста, так как она не учитывает характерную для каждого случая саморегуляцию. Она игнорирует также конституцию или возможную структуру яйца, которая может быть важным инициативным агентом, предшествующим {233} этим более поздним стадиям. Это строение должно быть теперь рассмотрено.
Может быть и окажется возможным дать чисто формальное объяснение, каким образом очень простая организация яйца может послужить основой для организации много более сложной, которая из него развивается. Но такая попытка в настоящее время неизбежно будет несовершенной и спекулятивной. Например, вокруг своей полярной оси вещество яйца имеет заметно слоистое строение. Можно предположить, что плоскость билатеральной симметрии обусловливается точкой проникновения сперматозоида. Дробление разделяет эти вещества по клеткам, причем одни содержат больше различных веществ, а другие — меньше. Можно предполагать, что эти различия в веществах влияют на последующее развитие отдельных областей яйца. Однако очевидно, что такая картина недостаточна для того, чтобы охватить все другие известные в настоящее время факты, касающиеся развития, ибо яйцо может быть разделено на куски, но полное развитие все-таки имеет место. Если постулировать такого рода постоянную организацию яйца, которая дает полностью организованного эмбриона, то половина ее должна дать только половину эмбриона. По-видимому, строгая прелокализация недостаточна для того, чтобы объяснить все возможности яйца. Постулируемая организация должна быть более подвижной или гибкой и способной к вторичным приведениям к норме. Поэтому некоторые эмбриологи приняли обыкновение говорить о таинственной самоорганизующей силе в яйце, не пытаясь установить, какого рода сила способна вызывать такие явления. Проблема этим только тормозится. Немного поможет делу также приписывание такой воображаемой организации особому молекулярному строению яйца. Никто не станет отрицать, что в яйце может быть определенное расположение молекул, но, пока эти идеи не смогут быть экспериментально проверенными и доказано их значение как в регулятивных процессах, протекающих в яйце, так и при регенерации у взрослых организмов, это допущение не принесет много пользы. Если клетки считать за единицы развития, то имеются данные, показывающие, что образование многих органов, повидимому, приводит целые группы клеток в координированные движения. Игнорируются ли границы клеток в этой организации? Нельзя упускать из вида, что во многих случаях, эти границы резко видны. {234}
Центрифугирование яиц показало, что по крайней мере их видимые вещества не являются органообразующими и так как вышеприведенная аргументация основана на их слоистости и перемещениях, то некоторые эмбриологи прибегли к допущению, что организация существует в «основной субстанции» яйца, т. е. в той прозрачной протоплазме, в которой мы можем видеть отсутствие диференциации. Такая попытка разрешения вопроса есть скорее отговорка, а не разъяснение проблемы. Она просто отодвигает проблему в бесконечность и ничего не дает для нашего познания. Если в основном веществе и существует такая организация, то мы можем, не нарушая ее, проводить сквозь нее желток, жиры и пигменты. Мы можем разрезать ее на куски и тем не менее получить новое целое из каждой части. Можно сказать, конечно, что каждый кусок реорганизует себя, но вопрос, как это происходит?
Несмотря на эти трудности, имеются данные, что в течение развития происходят более или менее необратимые прогрессивные изменения. Много раз было показано, что клетки и участки, еще не получившие видимой диференцировки, будучи изолированными, продолжают вести себя как части. Какова природа таких необратимых изменений? Ответ на этот вопрос будет, повидимому, так же важен для понимания нормального развития, как и для только что обсуждавшейся обратимости или реорганизации изолированных на ранних стадиях кусков.
Могут задать вопрос, что общего имеют строго физиологические изменения с видимыми изменениями, которые мы в совокупности называем развитием. Являются ли химические изменения следствием и непосредственным результатом физических изменений формы или, наоборот, они служат только для поддержания жизни организма, как это происходит у взрослого организма в течение его развития. В другой постановке последний вопрос может означать, что химические изменения заключаются главным образом в том, чтобы сделать вещества яйца пригодными для питания и снабдить яйцо энергией для физической стороны развития. Хотя такое разграничение может быть произвольным и, возможно, ошибочным, тем не менее оно, пожалуй, все же заслуживает краткого рассмотрения.
Растворение стенки зародышевого пузырька яйца, которое у одних яиц имеет место перед оплодотворением, у других же после него, казалось бы является химическим {235} явлением или, возможно, физическим переходом геля в золь. Когда это происходит после проникновения спермия, последний может выделять какое-либо вещество, которое в комбинации с веществами яйца может непосредственно вызывать эти изменения. Но когда стенка растворяется прежде, чем яйцо покинет яичник, то явление это представляется скорее автоматическим процессом, происходящим в самом яйце по достижении им определенной фазы своего развития. В этом отношении исчезновение оболочки ядра сравнимо с такого же рода изменением, происходящим перед делением каждого бластомера во время дробления. Растворение оболочки может быть вызвано чем-либо освобождающимся внутри ядра или может быть результатом изменений, происходящих во всей протоплазме.
Освобождение ядерного сока и смешение его с протоплазмой являются простым физическим изменением, которое, изменяя вязкость протоплазмы, ведет к ряду явлений, в числе которых находится также движение веретена к полюсу. Известно, что в это время конфигурация веществ яйца в смысле их видимого расположения в нем претерпевает изменения, что ведет к следующей ступени развития. Эти движения, чисто физические, происходят, повидимому, как следствие освобождения более жидкого ядерного сока.
Прободение поверхности яйца кончиком сперматозоида влечет за собой мгновенные изменения на поверхности яйца. Предполагалось, что само по себе прободение, изменяя натяжение оболочки, приводит к следующей фазе развития, но более вероятно, что происходит также какой-то локальный химический процесс, так как на поверхности яйца в точке прокола видны сильные изменения: происходит выделение воспринимающего бугорка и погружение в него сперматозоида.
При образовании воспринимающего бугорка на поверхности яйца приподнимается так называемая оболочка оплодотворения. О ее происхождении много спорили. Согласно некоторым наблюдателям, она присутствует перед оплодотворением и отстает от поверхности в результате скопления под ней жидкости; согласно другим, до оплодотворения эта оболочка отсутствует и образуется в момент оплодотворения в результате химического изменения на поверхности яйца. Было показано, что в это время проницаемость яйца изменена, и было предположено, что именно с этого и начинается развитие. С другой стороны, было обнаружено, {236} что оплодотворение и развитие яйца морского ежа могут происходить и без отставания оболочки. Очевидно поэтому, что изменения в яйце после оплодотворения могут происходить без образования оболочки или по крайней мере без ее отставания от поверхности яйца.
Эти начальные изменения дают много указаний на то, что они являются одновременно химическими и физическими реакциями, которые до этого момента не производят каких-либо резких изменений в яйце как целом. Так как отставание оболочки, за которым следует дробление яйца, может быть иногда вызвано или чисто химическими изменениями в составе морской воды путем таких физических приемов, как повышение температуры, или накалыванием, то, повидимому, правильно заключить, что эта реакция может быть объяснена в химических или физических терминах (или в тех и других) и не требует экстраординарного ультрафизического объяснения. Движение пронуклеуса яйца (после выделения полярных телец) и одновременное перемещение ядра сперматозоида по направлению к центру яйца могут быть смело приписаны физической реакции в результате набухания двух пронуклеусов, которое ведет их к общей точке. Повидимому, это является приспособлением, обусловленным увеличением размеров этих тел в жидкой среде, содержащей сферические тела различных величин. Другими словами, здесь, по-видимому, нет никакой нужды предполагать таинственные силы притяжения, действующие на расстоянии.
До сих пор изменения касались перемещений внутри самого яйца. Следующими изменениями являются видимые изменения формы всей системы как целого. Перед и во время деления яйца на две части на его поверхности и в подлежащем слое протоплазмы происходят хорошо видимые перемещения. Движения эти синхронны с образованием внутри яйца митотического веретена, которое, кроме других изменений, вызывает преобразование значительной части коллоидов яйца из золя в гель. Хотя мы знаем, что дробление яйца можно подавить химическими или физическими агентами и что в определенных пределах изменение скорости последовательных делений в зависимости от температуры приблизительно соответствует изменению скорости химических или физических реакций, тем не менее процессы, происходящие при клеточных делениях, слишком сложны для того, чтобы им можно {237} было дать чисто химико-физическое объяснение. Однако важно отметить, что почти не было попыток отнести эти факты к какой-либо области явлений, кроме как к механической.
Рост личинки и плода может рассматриваться как превращение запасов пищевых веществ в протоплазму. Значительная часть работ по химической эмбриологии основана на этом предположении. Физиология развития в этом смысле является в основном физиологией того же типа, как и физиология взрослого организма; различие заключается в значительной мере в том, что запасы веществ, необходимых для первых. стадий развития яйца, заключены в самом яйце, а для более поздних стадий поступают извне, как у взрослого организма.
Этот краткий обзор достаточен для того, чтобы показать, что эмбриолог вправе ожидать в будущем включения многих проблем развития в рамки законов физики и химии. Хотя эти физические явления и могут быть в большинстве случаев прослежены до изменений в отдельных клетках, но здесь имеется все же еще много не объясненного. Фактически большинство изменений, имеющих место в начале развития, происходит в пределах клетки. Яйцо есть развивающаяся клетка, и здесь, как нигде, мы имеем возможность непосредственно проследить физиологию клетки как индивидуума. Физиология взрослого животного имеет дело главным образом с группами клеток и взаимодействием их продуктов с остальным организмом. Нет оснований предполагать, что то, что имеет место в отдельных клетках органов, отлично от того, что происходит в отдельной яйцевой клетке...
Как я уже указывал, имеется интересная проблема, связанная с возможным взаимодействием между хроматином и протоплазмой клеток во время развития. Видимая диференцировка эмбриональных клеток происходит в протоплазме. Наиболее обычное генетическое предположение заключается в том, что гены в течение этого времени остаются неизменными. Однако допустимо, что по мере развития организма гены тоже все более и более усложняются или некоторым образом изменяются в соответствии с той частью протоплазмы, в которой они лежат и что эти изменения имеют обратное влияние на протоплазму. Можно возразить, что этот взгляд несовместим с тем, что путем изменения местоположения клеток, как это происходит в {238} опытах с прививками или при регенерации, клетки могут диференцироваться в другом направлении. Но это возражение не так серьезно, как кажется, если основное строение гена остается всегда тем же самым, а допускаемые усложнения или изменения в генах представляют собой явления того же порядка, что и те, которые происходят в протоплазме. Если протоплазма в новой внешней среде может менять свою диференцировку, не теряя своих основных свойств, то почему этим свойством не могут обладать также и гены? Этот вопрос находится, конечно, за пределами знаний, которыми мы располагаем в настоящее время, но как возможность нет нужды его отвергать. Ответа за или против такого предположения надо подождать до тех пор, пока экспериментальные исследования не смогут дать соответствующих доказательств.
| << | {239} | >> |
Вeer G. R. de, Growth, London, 1924.
Beer G. R., An introduction to experimental embryology, Oxford, 1926.
Beer G. R., Embryology and evolution, Oxford, 1930.
Brachet A., L'oeuf et les facteurs de l'ontogenese, Paris, 1917.
Dürken В., Lehrbuch der Experimentalzooiogie, Berlin, 1928.
Faure-Fremiet E., La cinetique du developpement, Paris, 1925.
Goldschmidt R., Physiologische Theorie der Vererbung, Berlin, 1927.
Gray I., A text book of experimental cytology, Cambrige, 1931.
Guyenot E., La variation et devolution, Paris, 1930.
Guyenot E., L'heredite, Paris, 1931.
Hоgben L., The nature of living matter, London, 1930.
Hogben L. a. Winton F. R., Comparative physiology, London, 1924.
Huxley I. S., Problems of relative growth, London, 1932.
Morgan Т. Н., Experimental embryology, New York, 1927.
Needham J., Chemical embryology, Cambridge, 1931.
Roux W., Entwicklungsmechanik der Organismen, Leipzig, 1895.
Russel E. S., The interpretation of development and Heredity, Oxford, 1930.
Schiep W., Die Determination der Primitiventwicklung, Leipzig, 1929.
Weismann A., Das Keimplasma, Jena, 1892.
Вильсон Э., Клетка и ее роль в развитии и наследственности, Биомедгиз, М., т. I, 1936, т. II (в печати).
Гексли Дж. и де Вер Г., Экспериментальная эмбриология, Биомедгиз, М.—Л., 1936.
Завадовский М. М., Внешние и внутренние факторы развития, Госиздат, М.—Л., 1928.
Завадовский М. М., Динамика развития организма, Медгиз, М.—Л., 1931.
Заварзин А. А., Краткое руководство по эмбриологии человека и позвоночных животных, Биомедгиз, Л., 1935.
Иванов П. П., Курс эмбриологии, Биомедгиз (в печати). {240}
Конклин Э. Г., Наследственность и среда, Госиздат, М.—Л., 1928.
Леб Ж., Организм как целое, Госиздат, М.—Л., 1926.
Лилли Ф. Р., Проблемы оплодотворения, Госиздат, М., 1925.
Морган Т., Структурные основы наследственности, «Госиздат, М.—Л., 1923.
Морган, Теория гена, «Сеятель», М., 1927.
Морган Т., Экспериментальные основы эволюции, Биомедгиз, М.—Л., 1936.
Филипченко Ю. А., Экспериментальная зоология, медгиз, Л.—М., 1932.
Шарп Л., Введение в цитологию, Биомедгиз, Л., (готовится)
1 Список составлен редакцией взамен чрезвычайно подробной и специальной библиографии оригинала.
| << | {241} | >> |
| << | {242} | >> |
Редактор Г. В и н б е р г. Техред. А. К а п е л ь к о. Зав. граф. ч. Е. С м е х о в. Зав. корректорской Л. Голицына. Ответ. за вып. в
типогр. П. М а р к е л о в.
Уполномоченный Главлита Б—15436 Биомедгиз 255. МД 86. Тираж 7200 Формат 82×109/32. Печ. л. 15,25. Знак, в печ. л. 35 100. Авт. л. 13,35 Сдано в тип. 25/VI 1936. Подп. к печ. 8/IV—1937 г. Заказ 656.
Цена 3 руб. Переплет 1 руб.
Набрано в 7 тип. «Искра революции» Мособлполиграфа, Москва, Арбат, Филипповский, 13. Отпечатано в 16 тип. треста «Полиграфкнига» Москва, Трехпрудный. 9.