
| {I} |
|
Природа и культура КНИГА ДЕВЯТНАДЦАТАЯ |
|
Проф. И. И. ШМАЛЬГАУЗЕН |
ПРОБЛЕМА
СМЕРТИ И БЕССМЕРТИЯ

|
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО МОСКВА * 1926 * ЛЕНИНГРАД |
| {II} |

| {III} |
Проблема бессмертия, выдвинутая в свое время главным образом Вейсманном, в последние годы в связи с усовершенствованием методов культивирования простейших и с введением метода тканевых культур вновь занимает одно из самых видных мест в биологической литературе.
С одной стороны, ряд экспериментов над некоторыми даже сложными животными (гидры, планарии) говорит в пользу возможности беспредельного продления жизни; с другой стороны, ясно, что явление смерти настолько связано с жизнью дифференцированных организмов, что проблема смерти является вместе с тем одним из основных вопросов о сущности жизненных процессов.
Мне хотелось ознакомить в доступной форме с накопившимися фактами и сделать попытку связать воедино кажущиеся противоречия.
Эта книга несколько задержалась в печати. За это время вышло довольно много новых работ, которые здесь уже не могли быть использованы. Некоторые дополнения я все же внес в виде примечаний позади текста (отмечены в тексте звездочкой). В общем же можно сказать, что новые факты не вносят изменений в наше понимание излагаемых вопросов. Цепь явлений, ведущих к старости и смерти, в общем совершенно ясна. В литературе различные авторы выдвигают лишь то или иное звено как руководящую причину явления. Таким образом, видят причину старческих изменений 1) в изнашивании организма (особенно нервной системы), 2) в накоплении продуктов обмена, отравляющих организм, 3) в возрастных изменениях внутрисекреторной системы, 4) в прогрессирующей дифференцировке, 5) в общем падении жизненной энергии как следствии возрастных изменений протоплазмы и, в частности, 6) в прекращении роста.
Все эти явления бесспорно являются, однако, лишь показателями возрастных изменений организма, а основная причина должна лежать глубже. {IV}
Мы в этой книге проводим мысль, что вся цепь явлений вытекает из истории развития организма как индивидуальности определенных размеров и формы. История развития любого организма есть история возникновения тормозящих влияний, которые удерживают процессы роста и ассимиляции в определенных рамках. Эти тормоза, ограничивая рост, ограничивают и возможность замены старого материала. Существование же возрастных изменений протоплазмы представляет собой, повидимому, прочно установленный факт.
И. Шмальгаузен.
14 января 1926 г.
Стр. |
||
1. | 1 | |
2. | 4 | |
3. | 11 | |
4. | 19 | |
5. | 26 | |
6. | 35 | |
7. | 40 | |
8. | 52 | |
9. | 63 | |
10. | 64 | |
11. | 74 | |
12. | 77 | |
13. | 80 | |
14. | 81 | |
15. | 85 | |
91 | ||
| {1} |
|
„Leben ist die schönste Erfindung der Natur und der Tod ist ihr Kunstgriff, viel Leben zu haben”. Goethe1). |
Жизнь — вечное движение, непрерывная смена веществ, структуры, формы. В этом постоянном потоке одна за другой выступают на сцену отдельные особи, проходят свой типичный жизненный цикл и умирают, уступая место другим. Существование каждой особи ограничено во времени. Возникая от других особей, живые организмы сами каждый раз заново воссоздают свою типичную структуру и сами в непрерывном обмене веществ и энергий поддерживают свое существование, пока, дав начало новым организмам, новой жизни, они не утеряют собственную, распавшись на те же безжизненные вещества, из которых они построили свое тело. Определенность жизненного цикла представляет собой одно из наиболее характерных свойств живых организмов. Зарождаясь в виде незначительного обыкновенно зачатка, как, например, одной клетки — яйца, организм при деятельном размножении клеток переходит в период интенсивного роста, связанного с развитием типической его формы и строения. Это — период юности, характеризующийся преобладанием процессов созидания (ассимиляции) над процессами разрушения (диссимиляции). Энергия роста зародыша в течение весьма короткого времени достигает очень значительной величины и затем еще на очень ранних стадиях развития довольно быстро падает.
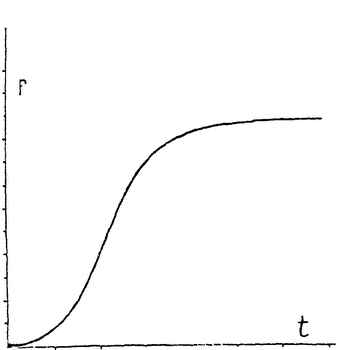
Кривая роста имеет довольно характерную форму буквы S, как это видно по прилагаемому чертежу (рис. 1), на котором по оси абсцисс отмечен возраст зародыша, а ординаты указывают его размеры. {2} Еще демонстративнее вторая кривая (рис. 2), указывающая интенсивность роста, вычисленную по процентам приращения веса в единицу времени. Здесь ясно видны быстрый подъем скорости роста и быстрое ее падение на весьма уже ранних стадиях развития. Указывают, что это падение энергии развития соответствует падению жизненной энергии клеток. Еще не родившийся организм вместе с приобретением жизни идет уже к смерти. Жизнь индивидуума во всех его фазах, начиная с самого зарождения, есть непрерывное умирание — некробиоз (Менерт). В этой парадоксальной фразе не мало истины. Но все же падение энергии клеточных делений еще не означает падения жизненной энергии вообще, оно
 |
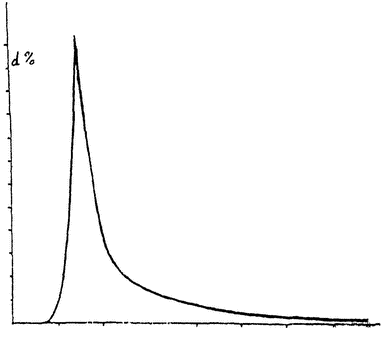 |
Рис. 1. Типичная кривая роста животного организма или отдельного органа. Абсцисса указывает возраст, а ордината — вес. |
Рис. 2. Типичная кривая интенсивности роста организма или отдельного органа. Абсцисса указывает возраст, а ордината — прирост в единицу времени, выраженный в процентах. |
означает лишь изменение характера жизнедеятельности. Процесса простого роста заменяются более сложными процессами развития со значительной затратой энергии на формообразование и клеточную дифференцировку.
Однако и процессы формообразования, как и рост, приходе к концу, и готовый организм переходит в период зрелости, который в общем характеризуется состоянием равновесия между процессами распада и восстановления. Наиболее важной особенностью зрелого организма является его способность к размножению. Как общее правило можно принять, что рост организмов является ограниченным, что вполне ясно выражено, например, у насекомые с метаморфозом, которые после вылупления из куколки уже вовсе не растут. То же самое наблюдается и у большинства наземных, {3} позвоночных, в том числе, как нам это известно из повседневного опыта, и у человека. В других случаях, однако, рост, постепенно, впрочем, угасая, продолжается в течение всей жизни. Это известно, например, для различных ракообразных, многих рыб, черепах.
В чем причина остановки роста? Несомненно, это прежде всего — результат прекращения клеточных делений. Теряют ли клетки с возрастом вообще способность к делению или в организме появляются какие-либо препятствия для клеточного размножения?
Экспериментальные исследования над культурами тканей вне организма показывают, что многие клетки пожизненно сохраняют способность к размножению, и с возрастом лишь падает скорость, с какой клеточные деления следуют друг за другом. Существование животных с постоянным ростом указывает на то же самое. Эксперименты с тканевыми культурами говорят в пользу того, что существуют в организме какие-то вещества (гормоны), стимулирующие клеточные деления (в тканевой жидкости зародышей, в ранах), и потому является весьма вероятным, что существуют и вещества, тормозящие таковые1. Поэтому следует думать, что клетки находятся в организме под влиянием гормонов, регулирующих их размножение. Во взрослом организме гормоны затормаживают клеточные деления, ведущие к увеличению размеров тела у животных с ограниченным ростом. Выделенные из организма клетки освобождаются от влияния тормозов и только тогда обнаруживают свою способность к клеточным делениям. То же самое иногда наблюдается и в организме, когда физиологическая изоляция известной части приводит к патологическим разращениям — постоянно растущим „злокачественным” опухолям (саркома, рак и др.). Нужно, однако, сказать, что не все клетки сохраняют способность к размножению. Некоторые особо высоко дифференцированные клетки, как мы увидим в дальнейшем, действительно окончательно теряют способность к размножению путем деления. Очевидно односторонняя их специализация несовместима с клеточными делениями. Это касается в особенности нервных клеток. Однако то же самое наблюдается нередко и в других органах, а иногда касается и всего организма, который в этом случае состоит из точно определенного числа клеток.
Однако рано или поздно животный организм переходит обыкновенно в третий период своей жизни — период старости, характеризующийся преобладанием процессов разрушения над процессами восстановления. Регрессивные изменения, дегенерация отдельных {4} органов расшатывают весь организм. Нарушается нормальная координация частей; правильное функционирование важнейших органов становится невозможным, и жизненные явления постепенно угасают. Наступает смерть. Таким образом старческая дегенерация входит в нормальный цикл особи, и смерть есть последнее звено в цепи жизненных явлений.
Смерть неразрывно связана с жизнью уже потому, что всякое проявление жизни возможно лишь путем уничтожения известного количества живого вещества. Смерть представляет собой как бы негативную сторону жизни. Без жизни нет смерти, и источником последней является сама жизнь.
Микроскопические животные организмы, известные под названием простейших, так же, как и сложные животные, обладают ограниченным ростом. Однако по достижении своих предельных размеров животное не умирает, а более или менее сложным путем
 |
Рис. 3. Деление амебы (Amoeba polypodia) по Ф. Шульце из Ланга. |
На рис. 3 показан процесс деления амебы. Как видно на рисунке, делению протоплазматического тела амебы предшествует деление ее ядра, которое вытягивается в длину, приобретает форму гантели и затем разрывается по средине. Чем дальше подвигается дело изучения простейших, тем более накопляется фактов, позволяющих провести сравнение между этим делением простейших и сложным, кариокинетическим делением клеток высших организмов. И здесь частицы важнейшей ядерной субстанции — хроматина — равномерно распределяются между ядрами дочерних особей. {5}
При делении более сложно построенных простейших — инфузорий — явление осложняется еще существованием органов, имеющихся в единственном числе и не могущих быть разделенными. В этом случае данный орган переходит к одной из дочерних особей, а в другой особи подобный же орган развивается заново. На рис. 4
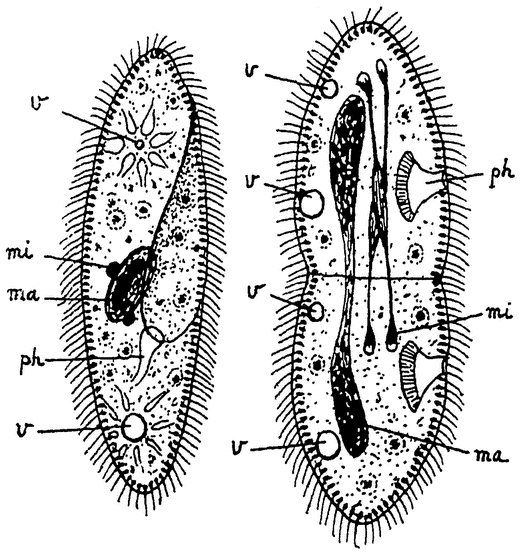 |
Рис. 4. Деление туфельки (Paramaecium aurelia). По Д. Третьякову. |
В этих процессах, как впервые отметили Вейсманн и Бючли, нет ничего похожего на смерть. Смерть есть прекращение жизни и сопровождается всегда появлением трупа. Здесь нет ни того, ни другого. Все продолжает жить, и ничто не погибает.
Таким образом простейшие бессмертны, или, точнее, потенциально бессмертны, так как у них нет естественной, физиологической смерти, между тем как при неблагоприятных условиях существования они, разумеется, гибнут массами.
При делении животного надвое, понятно, теряется индивидуальность материнской особи, — она как бы растворяется в дочерних. Объективно такая потеря индивидуальности со смертью не сравнима. В некоторых случаях, однако, трудно говорить даже и о потере индивидуальности. Некоторые простейшие размножаются путем обособления лишь незначительной части своего тела, путем так называемого почкования (особенно у сосущих ацинет). В этом случае материнская особь, очевидно, вполне сохраняет свою индивидуальность. Между таким почкованием и обычным делением надвое наблюдаются все переходы в виде неравных делений различного типа, и трудно указать, где индивидуальность материнской особи при этом теряется, где нет. {6}
Время от времени в культурах простейших наблюдаются чрезвычайно интересные явления, которые с полным основанием сравнивают с половым процессом у сложных животных. У простейших этот половой процесс наблюдается в виде временного соединения двух особей, так называемой конъюгации, или полного их слияния, что обозначают как копуляцию. И в том и в другом случаях наиболее интересными являются сложные преобразования в ядерном аппарате обеих особей.
Половой процесс наблюдается лишь после более или менее значительного периода размножения. К концу такого периода интенсивность размножения значительно падает, так же, как падает и вообще вся жизнедеятельность особей. Обыкновенно такие изменения (состояние „депрессии”) наступают сразу у большинства особей данной культуры и разрешаются затем целой эпидемией конъюгации.
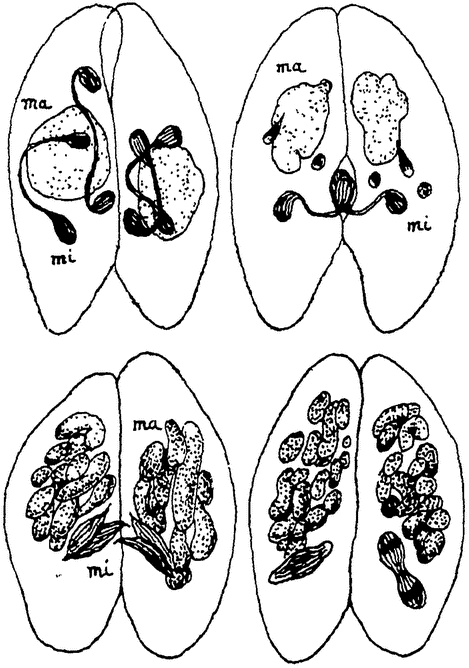 |
Рис. 5. Конъюгация туфельки (Раramaecium). По Р. Гертвигу. ma — макронуклеус, mi — микронуклеус |
Ход последней лучше всего изучен у ресничных инфузорий и в особенности у упомянутой уже парамеции. Две парамеции прикладываются друг к другу своими ротовыми отверстиями и слипаются. Вслед за этим наступают резкие изменения в ядерном аппарате (рис. 5). У парамеции, как и у других инфузорий,— два ядра: большое, вегетативное ядро — macronucleus, играющее важнейшую роль в процессах обмена веществ, и другое, малое, генеративное ядро — micronucleus, которое, как мы сейчас увидим, служит для восстановления всего ядерного аппарата при половом процессе. Большое ядро в обеих особях перерождается, распадается и бесследно погибает. Малое ядро делится типичным образом дважды подряд и дает таким образом начало четырем ядрам в каждой особи. Из этих четырех ядер три распадаются, а четвертое делится еще раз на два ядра, из которых одно остается на месте, а другое перемещается в конъюгирующую особь. Таким образом происходит обмен ядрами между обеими особями. {7} Каждая из них сохраняет одно свое ядро и получает второе от другой особи. Эти ядра сливаются и путем деления воссоздают затем весь нормальный ядерный аппарат, т.-е. как micronucleus, так и macronucleus в каждой особи, после чего конъюганты расходятся и вновь приступают к размножению посредством деления.
В обмене ядрами мы видим наиболее существенный момент полового процесса, сравнимый с оплодотворением у высших животных (амфимиксис). Так как конъюгация наблюдается в культурах обычно при известном недостатке пищи, обнаруживающемся в результате чрезмерного размножения инфузорий, то явилась мысль воспрепятствовать их конъюгации путем постоянного обильного снабжения культур свежим питательным материалом. Такие опыты действительно удались сначала Мопа, затем Калкинсу и другим исследователям. Конъюгацию удалось предотвратить, но понижение жизнедеятельности — состояние „депрессии” целой культуры все же периодически наступало. Путем обновления питательной среды, а затем путем внесения различных экстрактов, действовавших в качестве химических раздражителей, удавалось прерывать это состояние и возбудить инфузорий к новому подъему жизнедеятельности и к интенсивному размножению. Однако все эти меры оказывались действительными лишь до поры до времени. В конце концов наступали полная дегенерация и гибель всей культуры (у Калкинса на 742-м поколении). Эти наблюдения вновь выдвинули вопрос о физиологической смерти у простейших. Мопа и другие авторы стали видеть в состоянии депрессии явления старческого вырождения, которое и у простейших ведет к смерти, если только животное не избегнет ее благодаря омолаживающему влиянию полового процесса.
Таким образом возникла новая постановка всей проблемы, и Р. Гертвигом и другими авторами развито замечательно красивое сравнение жизненного процесса сложного животного с полным циклом размножения простейшего. Как инфузория после конъюгации приступает к деятельному размножению, интенсивность которого затем после известного числа поколений падает, и все особи при явлениях депрессии приближайся к физиологической смерти, так и у сложного животного оплодотворенная яйцеклетка размножается путем простого деления, сначала чрезвычайно интенсивно, а затем все медленнее; жизнедеятельность этих клеток, составляющих целый организм сложного животного, падает, и они при явлениях старческого перерождения приближаются к смерти. {8} Продукты клеточных делений в этом случае лишь остаются во взаимной связи, а не расходятся для ведения самостоятельной жизни, как у большинства простейших. Имеется, однако, еще одно существенное различие. У простейших все особи последней генерации способны к омоложению в результате полового процесса, между тем как в организме высших животных эту способность сохраняют одни только половые клетки.
Но ведет ли половой процесс, и в частности конъюгация, действительно к омоложению организма? Это является в высшей степени сомнительным уже потому, что далеко не у всех простейших половой процесс является столь обязательным, и даже у парамеций, по исследованиям Дженнингса, имеются различные расы, отличающиеся по своей потребности в конъюгации. Одни расы конъюгируют часто, другие редко, а у третьих и вовсе не наблюдалось конъюгации в течение 2—3 лет. Наконец, Дженнингс произвел еще следующий опыт: он разделял соединившихся для конъюгации парамеций и культивировал их дальше. Оказалось, что жизнеспособность этих культур была даже выше контрольных, где конъюгация была доведена до конца.
Позднейшие эксперименты Вудрёф и Метальникова проливают новый свет на весь этот вопрос. Уже Мопа и Калкинс при своих опытах пришли к необходимости постоянного изолирования минимального числа особей в сравнительно большом количестве жидкости. В противном случае состояния депрессии наступают очень быстро вследствие накопления продуктов обмена, что и приводит к гибели всю культуру. Вудрёф применяла собственно тот же метод ведения культур, но довела его до совершенства путем ежедневного изолирования одной особи (Paramaecium aurelia делится в среднем 1—2 раза в сутки) в 5 каплях жидкости (сенной отвар). Возможность конъюгации была при этом методе совершенно исключена. Таким образом Вудрёф удалось с мая 1907 года до декабря 1920 года получить 8400 поколений, поддерживая культуру в продолжение 131/2 лет. Если бы возможно было поддерживать существование всех получаемых особей, то эта культура привела бы к образованию невероятно большого числа особей (28400) с массой протоплазмы, неизмеримо превышающей массу земного шара. По всей вероятности эта культура поддерживается и в настоящее время. Такой продолжительный срок и колоссальное количество полученных поколений дают нам уже полное право вывести заключение, что при благоприятных условиях существования размножение {9} простейших путем деления может продолжаться безгранично и без вмешательства процесса конъюгации. Дегенерация не наступает, и инфузории вполне сохраняют свою жизнеспособность. Конъюгация, следовательно, не является омолаживающим фактором, а имеет какое-то иное биологическое значение. Такие же культуры велись и у нас в России проф. Метальниковым и в течение 1910—1920 годов привели к образованию путем деления 4 000 поколений.
Точно также и Дофлейн в течение многих лет культивировал патогенных трипанозом на постоянно сменяемых питательных средах без всяких признаков дегенерации, хотя размножение путем деления совершенно не прерывалось половым процессом. То же самое достигалось и с амебами (Эрдманн, Дофлейн) и с биченосцами (Дофлейн, Гартманн). Наконец Гартманн культивировал также и известную колониальную Eudorina elegans в течение более 5 лет при исключительно бесполом размножении, без всяких признаков депрессии (свыше 1400 индивидуальных поколений).
Таким образом ясно, что конъюгация сама по себе не является омолаживающим фактором. Однако с конъюгацией теснейшим образом связан другой процесс, который, очевидно, и направлен к обновлению всего организма, это — та сложная перестройка ядерного аппарата, которая является подгоговкой к обмену ядрами при конъюгации. Это видно из позднейших наблюдений.
Уже Калкинс отметил при своих культурах парамеций периодические изменения скорости делений, и то же самое подтверждено Вудрёф. По новейшим исследованиям Вудрёф и Эрдманн, эти ритмические колебания находятся в связи со сложными изменениями в ядерном аппарате. Последние вполне сходны с теми, что наблюдаются при конъюгации. После известного числа простых делений большое, вегетативное, ядро парамеции теряет правильность своих очертаний и распадается (рис. 6). Малые, генеративные, ядра (у Paramaecium aurelia их два) увеличиваются в размерах и затем делятся дважды, при чем из четырех продуктов деления три погибают, а за счет четвертого, оставшегося, ядра восстанавливается затем Заново весь ядерный аппарат (1 вегетативное ядро и 2 генеративных). Здесь происходит, следовательно, вся подготовка к половому процессу, но нет самого существенного при конъюгации — обмена ядрами двух особей (амфимиксиса). С полным основанием это явление рассматривают как партеногенез, т.-е. девственное размножение. {10}
Описанные преобразования ядерного аппарата совпадают с падением интенсивности размножения, а после того как они закончены наблюдается значительное ускорение делений, а также и вообще повышение жизнедеятельности.
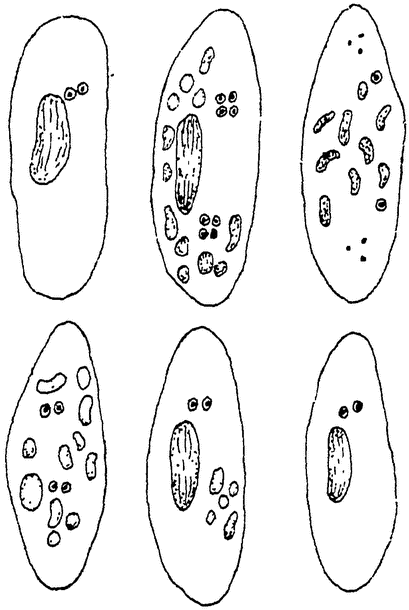 |
Рис. 6. Перестройка ядерного аппарата у туфельки (Paramaecium) по Вудрёф и Эрдманн из Коршельта. Микронуклеусы отмечены точкой в кружочке. На первых трех стадиях видны полный распад макронуклеуса, деление микронуклеусов и гибель трех его четвертей. На последних трех стадиях — восстановление, всего ядерного аппарата из оставшейся четвертой части обоих микронуклеусов. |
Таким образом перестройка ядерного аппарата, очевидно, представляет собой процесс, ведущий к омоложению всего организма. Ясно, что и при конъюгации не обмен ядрами, не амфимиксис ведет к омоложению, а предшествующее ему характерное обновление ядерного аппарата.
Если и видеть в явлениях депрессии у простейших признаки старческого вырождения цикла, выражающиеся в ряду бесполых поколений в понижении жизнедеятельности и падении интенсивности деления, то все же простейшие бессмертны, так как они обладают механизмом, автоматически вызывающим в этом случае их омоложение. Явления старости имеются, но смерть целого не наступает благодаря периодическому омоложению.
Частичная смерть простейших. Инфузории обязаны своим бессмертием периодическому обновлению ядерного аппарата. При этом совершенно распадается и гибнет прежде всего большое, вегетативное ядро, которое является главным регулятором всех жизненных процессов, но затем погибает также еще и большая часть малого, генеративного ядра. Таким образом значительная часть тела инфузорий имеет ограниченное существование. Так как это как раз центральный регуляторный аппарат, без которого жизненные явления невозможны, то, очевидно, здесь имеется налицо настоящее изнашивание. Гибель вегетативного ядра можно рассматривать как результат старческой дегенерации, ведущей к частичной смерти. Подобные явления наблюдаются и в других случаях. Так, например, у солнечников при энцистировании гибнет большая часть {11} ядерного аппарата. Еще гораздо яснее видна частичная смерть у некоторых других простейших. Большой морской биченосец-ночесветка (Noctiluca) размножается путем множественного почкования (рис. 7). Каждая почечка дает начало подвижной споре. После образования спор остается, однако, значительная часть тела, которая оказывается нежизнеспособной и скоро гибнет. Это уже настоящий труп, и притом образование его связано с прекращением каждой индивидуальной жизни, подобно тому как и у сложных животных.
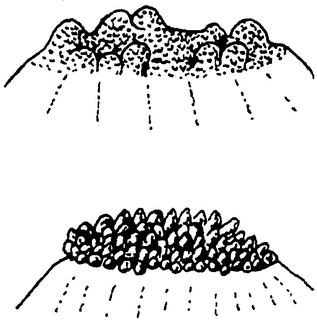 |
Рис. 7. Почкование ночесветки (Noctiluca) из Р. Гертвига. Передний конец тела с образующимися зооспорами. |
То же самое наблюдается и во многих других случаях при единовременном образовании большого числа подвижных спор. Всегда множественное деление и почкование сопровождаются появлением более или менее значительного остаточного тела. В особенности характерен такой способ размножения для споровиков (Sporozoa).
Наконец у многих простейших некоторые органеллы отбрасываются и затем вновь восстанавливаются регулярно при каждом делении. Таким путем возобновляются обыкновенно более сложно построенные органы движения — жгутики, щетинки, а также вакуоли, рот и другие органеллы. Таким образом все же у простейших некоторые части тела окончательно изнашиваются и обладают ограниченным существованием. Это касается в особенности наиболее специализированных, частей — ядерного аппарата и более сложных органоидов.
Всякая жизнедеятельность связана с тратой живого вещества. Последнее вначале успешно восстанавливается, но с течением времени это восстановление становится все менее совершенным, жизнедеятельность падает, и таким образом проявляется более или менее заметное изнашивание.
Простейшие животные обладают многими средствами борьбы с этим изнашиванием. Прежде всего уже при каждом делении происходит известная реорганизация ядерного аппарата (на котором изнашивание всего более сказывается), и, кроме того, иногда отбрасываются и затем восстанавливаются заново те или иные {12} дифференцированные части тела. При наступлении неблагоприятных условий существования у простейших может наступить энцистирование, т.-е. образование плотной оболочки, под защитой которой животное может продолжительное время пребывать в состоянии покоя. Энцистирование также связано со значительной реорганизацией особи, которая при восстановлении благоприятных условий вылупляется совершенно обновленной. Наконец у простейших наблюдаются еще периодические глубокие преобразования, связанные с гибелью значительной части ядерного аппарата и других высоко дифференцированных частей и ведущие к полному обновлению всей организации. Этот процесс иногда связан со взаимным обменом и установлением новой комбинации ядерных веществ двух особей при явлении конъюгации (амфимиксис), но возможен и без этого (партеногенез). Благодаря всему сказанному у одиночных простейших обычно нет физиологического умирания целой особи.
Однако у колониальных форм иногда намечаются уже иные соотношения. Прежде всего рост колоний обыкновенно ограничивается известными пределами, и в связи с этим устанавливается нередко строгое фиксирование определенного числа делений, ведущих к образованию колоний. У различных сувоек наблюдаются древовидные колонии с определенной системой ветвления. Некоторые виды обладают, повидимому, неограниченным ростом, между тем как другие имеют строго определенные размеры, при чем в связи с системой ветвлений данного вида некоторые особи, занимающие известное положение в системе, очень рано теряют способность размножения посредством деления. Лучше изучены некоторые колониальные биченосцы. У Gonium и Pandorina уже 4 последовательных деления приводят к образованию нормальной колонии из 16 особей. При вегетативном размножении отдельные особи раздвигаются шире, и затем каждая приступает к повторному делению и дает начало новой, дочерней колонии в 16 особей. Колония может также распасться на отдельные половые особи — гаметы, которые после попарной копуляции и образования зиготы начинают сызнова процесс бесполого размножения. У Eudorina 5 последовательных делений приводят к образованию колонии в 32 особи. Как и у предшествующих форм, каждая особь может вегетативным путем дать начало новой колонии. С наступлением половой зрелости в этих колониях наблюдается ясная половая дифференцировка — в одних, женских колониях, сохраняется нормальное число 32 половых особей (макрогамет), между тем как {13} в мужских колониях каждая особь распадается на целый пучок небольших удлиненных микрогамет. Как мужские, так и женские гаметы снабжены и жгутиками и глазными пятнами. И в этом случае, следовательно, все особи колонии могут приобрести характер половых особей, которые притом существенно не отличаются от обычных вегетативных особей. Половой процесс,, однако, и в этом случае так же мало необходим для поддержания жизни вида, как и у одиночных простейших, и мы видим, что Гартманну удалось поддерживать, повидимому, бессмертную культуру Eudorina elegans при непрерывном вегетативном размножении, вполне исключив половое воспроизведение.
У других представителей колониальных биченосцев — известного рода Volvox — дифференцировка и специализация идут гораздо
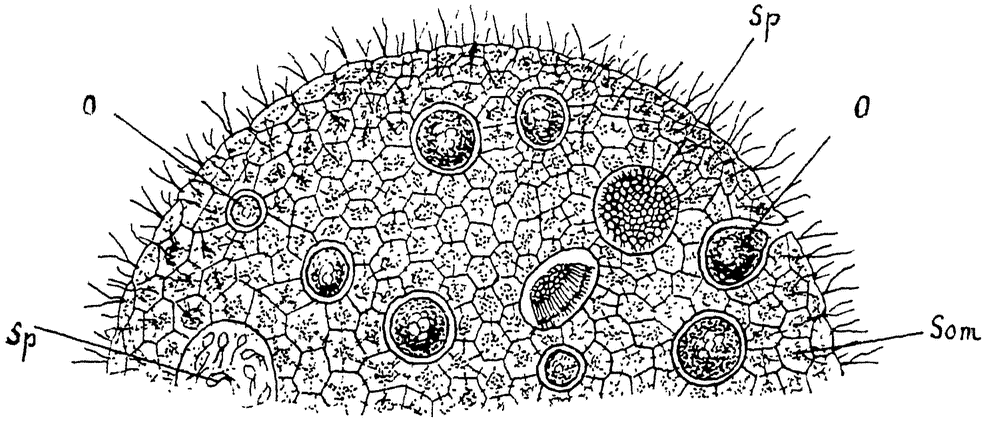 |
Рис. 8. Колония Volvox globator. По Лангу из Гертвига: Som — соматические особи, О — женские половые (яйца), Sp — мужские половые особи (сперматозоиды). |
дальше. Это сравнительно огромные колонии, состоящие из большого (многих тысяч) числа особей, теснейшим образом, подобно клеткам сложных животных, связанных между собой протоплазматическими мостиками (рис. 8). Отдельные особи снабжены, как и в ранее описанных колониях, жгутиками и глазными пятнами и правильно распределены по поверхности шарообразной колонии.
Самое интересное в этих крупных колониях — глубокая дифференцировка на соматические особи, несущие все обычные жизненные функции — питания, движения, раздражимости, и половые особи, являющиеся в виде очень крупных макрогамет и мелких веретеноидных микрогамет, развивающихся значительными пучками. Макрогаметы лишены органов движения и имеют шаровидную {14} форму, живо напоминая собой яйца сложных животных. Микрогаметы очень подвижны и вполне сравнимы со сперматозоидами. В общем рост этих колоний также ограничен известными пределами. Способность отдельных особей к делению угасает. Исключение составляют лишь некоторые особи, которые путем возобновления многократного интенсивного деления дают начало дочерним колониям („партеногонидии”), лежащим внутри старой. Возможно, что в случае такого вегетативного размножения возобновлению процесса деления предшествует перестройка ядерного аппарата, подобно тому как это мы видели у парамеций1).
После образования половых элементов и партеногонидий колония погибает. Пример Volvox поучителен в особенности потому, что, с одной стороны, мы имеем действительно лишь колонию типичнейших биченосцев, с другой стороны, отдельные ее особи связаны между собой так же тесно, как и у сложных (многоклеточных) животных, и, как и у последних, существует ясная дифференцировка на соматические и половые элементы.
Половые элементы служат для воспроизведения следующих поколений; подобно любой особи одиночных простейших они сохранили потенциальное бессмертие. Остальные особи колонии, составляющие главную массу его тела (сомы), специализировались для несения обычных жизненных функций; их способность к делению быстро угасает, и в большинстве случаев, очевидно, утрачивается и способность к обновлению ядерного аппарата; они утратили свое бессмертие. Так как эти особи представляет собою главную массу колонии, то их гибель вполне сравнима со смертью сложных животных, для которых столь характерно образование объемистого трупа.
С чем связано появление смерти у колониальной формы, подобной Volvox? Смерть сложного организма обычно объясняют специализацией его сомы. С физиологической точки зрения это может быть и так. Однако для биолога этим вопрос не исчерпывается. Ключ должен быть найден путем изучения колониальных биченосцев, где мы впервые наблюдаем появление смерти. Здесь специализация сомы вряд ли играет столь существенную роль, так как эта специализация выражается лишь в выпадении одной из функций — полового воспроизведения. {15}
Кроме этой специализации мы видим у всех колониальных форм только еще одно новое обстоятельство — появление новой, высшей индивидуальности определенной формы и размеров. Вот эта определенность формы и объема колонии неизбежно связана с ограничением роста, т.-е. координированным прекращением делений отдельных особей. Число последовательных делений может быть ограничено 4—5, как мы это видим в колониях Gonium, Pandorina и Eudorina. Ограничение это определяется какими-то специфическими внутренними причинами, затормаживающими рост и деление на строго определенной стадии. Биологическое значение определенности формы и размеров понятно. С этим связано ограничение делений, а следовательно, и раннее выпадение обновления ядерного аппарата, характерного для всякого „непрямого” деления.
Возможность преобразования любой особи в половую показывает, однако, что способность глубокой перестройки ядерного аппарата, как она наблюдается при половом процессе, окончательно еще не утеряна. Следует думать, что и вегетативному размножению предшествует обновление ядерного аппарата. У Volvox, однако, большинство особей несомненно теряет способность обновления ядерного аппарата; они сохраняются неизменными в качестве элементов сомы, и таким образом обеспечивается более продолжительное существование высшей индивидуальности — всей колонии в целом. При отсутствии делений во взрослой колонии дифференцированные части, и в особенности регуляторный аппарат отдельных особей, неизбежно изнашиваются, и в конце концов вся колония приближается к естественному концу — физиологической смерти.
Колония Volvox при воспроизведении не распадается целиком на составляющие ее элементы, как у Gonium, Pandorina и Eudorina; жизнь ее как целого сохраняется довольно продолжительное время. Целость колонии сохраняется именно благодаря существованию невоспроизводительных — соматических — особей. Благодаря последним жизнь колонии как особи высшего порядка оказывается гораздо более продолжительной, но зато благодаря им же все, что не использовано на воспроизведение новых колоний, осуждено на естественную смерть.
Таким образом возникновение физиологической смерти теснейшим образом связано с появлением высшей индивидуальности ограниченных размеров при условии дифференцировки на воспроизводительные особи и соматические. Отдельные особи утрачивают способности размножения в связи с ограниченностью роста колонии. {16} Обособление сомы имеет свое биологическое значение в продлении жизни высшей индивидуальности — колонии, как целого. Оно связано с ограничением роста, а следовательно с ограничением ассимиляции и утратой способности обновления ядерного аппарата (это привело бы к размножению и распаду колонии), а в результате отсутствия реорганизации наступают старческая дегенерация и смерть.
Все сложные животные обладают в конце концов ограниченным
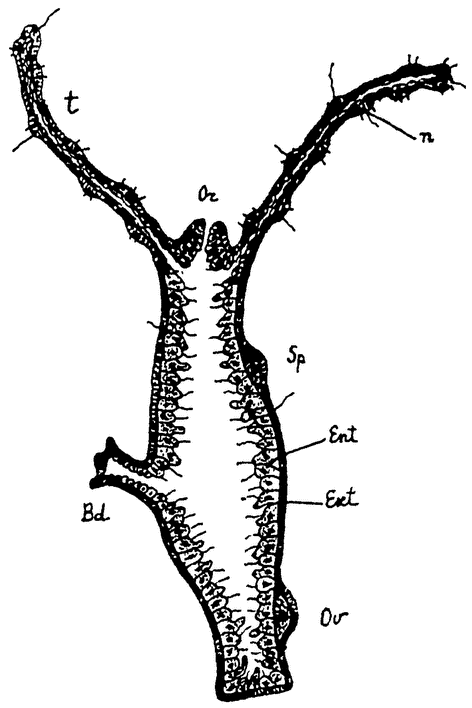 |
Рис. 9. Схематический разрез через тело гидры. Bd — почка, Ekt — эктодерма, Ent — энтодерма, n — стрекательные клетки, Or — рот, Ov — яичник с одним зрелым яйцом, Sp семенник, t — щупальце. |
Примером сравнительно еще низко стоящего сложного животного мы возьмем обыкновенного пресноводного полипа — гидру (рис. 9). Как колония Volvox развивается путем многократного деления одной оплодотворенной макрогаметы, так и любое сложное животное, в том числе и гидра, развивается путем многократного деления одной оплодотворенной яйцеклетки. Из последней развивается полый пузырек (в общем напоминающий Volvox), состоящий сначала из одного, а затем из двух слоев продуктов этого деления — клеток. Пузырек принимает удлиненную цилиндрическую форму. Одним своим концом он прикрепляется к субстрату, а на другом конце появляется „ротовое” отверстие, ведущее во внутреннюю полость. Вокруг ротового отверстия образуется некоторое число (6—8) длинных полых выростов — щупальцев.
Рост гидры ограничен, и форма ее определенна, однако до известной только степени. При обильном питании вырастают более крупные гидры, с бóльшим числом щупалец, чем при скудном питании. {17}
Тело гидры состоит только из двух слоев клеток: наружного слоя, образующего общие покровы,— эктодермы и внутреннего слоя, выстилающего внутреннюю, пищеварительную полость, — энтодермы. Между клетками наблюдается, однако, уже очень значительная дифференцировка. Клетки энтодермы, правда, довольно однообразны; они снабжены жгутиками и несут главным образом только функции питания. Среди клеток эктодермы находятся кроме обычных цилиндрических клеток эпителия особые защитные стрекательные клетки, чувствующие клетки, а несколько глубже — индифферентные, так называемые интерстициальные клетки, и более крупные, нервные. В глубине эктодермы лежат также мускульные отростки эпителиальных клеток, несущие функции мускульной ткани (подобные же мускульные отростки имеются и на энтодермальных клетках). Наконец в эктодерме же развиваются и половые клетки — крупные яйца по одиночке у проксимального конца тела и мелкие сперматозоиды, расположенные значительными группами недалеко под венчиком щупальцев. Гидра сохранила способность и вегетативного размножения. При хорошем питании на ее теле в различных местах могут образоваться полые выросты, которые развиваются в новых гидр, отделяющихся от тела матери и прикрепляющихся затем самостоятельно к какому-либо субстрату.
При хорошем питании, следовательно, сдерживающее влияние целого организма местами прекращается; возобновляются клеточные деления, которые приводят к образованию новой типической структуры — новой особи. Способность к воспроизведению сохранилась постольку, поскольку это возможно без нарушения целости всего индивидуума. Последний многократно воспроизводит дочерние особи, обладает относительно чрезвычайно продолжительной жизнью (до одного года, а иногда и дольше), но бессменно функционирующие соматические его клетки, особенно, вероятно, нервные, без обновления ядерного аппарата в конце концов все же изнашиваются и ведут организм к смерти.
Если мы теперь перейдем к рассмотрению организации какого-либо из высших животных — моллюска, насекомого или позвоночного, — то найдем картину неизмеримо более сложной дифференцировки. Отдельные части тела специализировались в различных направлениях и являются в виде органов того или иного назначения. Каждый орган представляет собою сложное образование — соединение различных тканей, в свою очередь состоящих из множества клеток того или иного типа. Но что самое важное в этой сложности, {18} это — точная определенность всей организации. Не только общая форма животного, число, распределение и состав его органов вполне предопределены, но даже тончайшая, интимная структура отдельных частей. Нередко даже число, расположение, форма и связи клеток важнейших органов, как, например, нервной системы, строго фиксированы, а иногда и все органы, а следовательно и все животное, состоят из точно определенного числа клеток.
Весь этот организм развивается, подобно прежде рассмотренным, путем многочисленных клеточных делений из одной единственной яйцеклетки. Вначале равномерные деления, как и у гидры, приводят к образованию полого шара, но затем неравномерный рост приводит к изменению общей формы, образованию складок, впячиваний, плотных разращений и т. д. Путем обособления областей разной скорости роста и особой дифференцировки образуются зачатки будущих органов. Здесь, следовательно, имеется точно действующий внутренний регуляторный аппарат, который прекращает клеточные деления на строго определенной стадии.
При такой определенности строения возможен еще только гармонический рост всего организма (хотя и он рано прекращается, так как и величина животного ограничена известными пределами), но совершенно исключается возможность общего возобновления клеточных делений в той или иной части тела, так как она нарушила бы, хотя бы и на время, общую гармонию организации. Очевидно с этим связана и полная утрата той способности вегетативного размножения (путем деления или почкования), которая, как мы видели на примере гидры, характерна для многих более низко организованных животных.
Только одна группа клеток не специализируется для несения обыденных функций, не участвует в общей жизни организма и сохраняет в неприкосновенности свою способность к воспроизведению. Это — половые клетки. В процессе индивидуального развития и они принимают участие, и они питаются, растут и размножаются путем деления, и они живут, но лишь своей обособленной жизнью. Изнашивание и падение способности вегетативного размножения наблюдаются, очевидно, и здесь. Но эти воспроизводительные элементы сохраняют зато полностью способность к обновлению своей организации путем перестройки ядерного аппарата — явление, известное как созревание половых продуктов.
Специализация соматических клеток высших животных связана с более совершенным выполнением определенных функций, а при {19} точной координации деятельности отдельных клеток и различных органов сложного организма она обеспечивает наилучшие условия для их существования, и таким образом не должно удивлять, что эта- специализация приводит к значительному увеличению продолжительности жизни отдельных клеток. У высших животных эта продолжительность жизни более специализированных и потому не сменяемых клеток (например нервной системы) определяется жизнью всей особи, т.-е. измеряется годами, десятками, а иногда и сотнями лет. Стойкость соматических клеток значительно возросла благодаря их специализации, при чем наиболее стойки всегда наиболее специализированные, нервные и мускульные, клетки, как это доказывается экспериментами голодания у планарий и других животных (Е. Шульц и др.).
Но бессменно функционирующие клетки, не обновляемые ни путем деления, ни путем какой-либо иной перестройки ядерного аппарата, в конце концов изнашиваются. Точная координация функций различных частей организма нарушается, наступают явления старости, заканчивающиеся прекращением жизнедеятельности всего организма. Наступающая смерть есть результат образования высшей индивидуальности. Смерть является платой за продление жизни особи как гармонического и стойкого целого с высоко развитой индивидуальностью.
Мы видели, что возникновение смерти связано с прогрессивным развитием стойкой высшей индивидуальности ограниченных размеров и строго определенной формы с вполне гармоничным соотношением частей. Строгое ограничение числа клеточных делений и точная координация их хода в отдельных частях организма предполагают существование особого внутреннего аппарата, определяющего эти соотношения. Сущность этого аппарата до сих пор не выяснена, но, основываясь на изучении развития животных, можно уже указать, что осуществление формы организма достигается путем строгой детерминации, а осуществленная уже форма сохраняется путем постоянной регуляции.
Под детерминацией мы понимаем существование известной внутренней структуры зачатка, благодаря которой из данной части может развиться только строго определенная система. Если яйцо, а иногда и первые продукты его деления, обладает способностью {20} воспроизвести весь организм, то по мере дальнейшего развития „проспективная” способность отдельных клеток все более ограничивается лишь известной областью, пока, наконец, окончательно не определится их назначение в качестве мускульной, железистой или определенной нервной клетки и т. д.
Назначение отдельных частей определяется вначале их соотношением, но в дальнейшем они развиваются путем „самодифференцирования”, т.-е. независимо от соотношения с другими частями.
Обычно говорят о детерминированном развитии только тогда, когда оно проявляется очень рано, когда предопределено назначение уже первых продуктов деления яйцеклетки. В этом случае уже прото-
 |
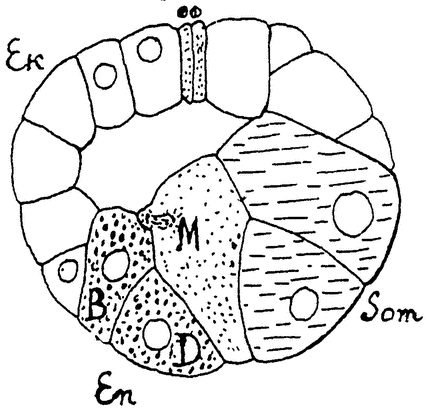 |
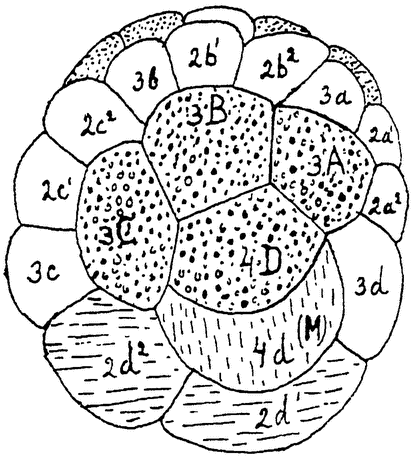
Рис. 10. Спиральное дробление кольчатого червя (Arenicola) по Чайльду из Коршельта и Гейдера. Кружочками отмечены энтобласты (будущая энтодерма), вертикальной штриховкой — мезобласт (будущая мезодерма), горизонтальной штриховкой — соматобласты (будущая эктодерма туловища червя), пунктиром — трохобласты будущее предротовое ресничное кольцо личинки), белыми оставлены остальные эктобласты (будущая эктодерма головной части). |
Рис. 11. Сагиттальный разрез через зародыш кольчатого червя (Amphitrite) на стадии бластулы. По Виду из Давыдова. Еn — энтобласты, М — мезобласты, Som — соматобласты, Ek — остальные эктобласты. |
плазма яйца всегда оказывается дифференцированной, и первые же продукты деления отличаются друг от друга по своему составу. Здесь дифференцировка начинается раньше, чем развитие зародыша, и даже раньше, чем произойдет оплодотворение,—уже при созревании яйца в яичнике. Примером такого детерминированного развития могут служить моллюски и кольчатые черви (рис. 10, 11). Яйцо в этих случаях делится по строго определенному, так называемому спиральному, типу, на клетки различного состава и назначения. Каждая клетка дает начало только определенным органам, их проспективная способность строго ограничена. Если удалить известную {21} часть яйца, то зародыш разовьется с соответствующими дефектами.
Однако и в других случаях, когда детерминация на ранних стадиях не выражена, когда первые продукты деления совершенно равнозначущи и могут замещать друг друга, с началом дифференцировки, с намечающимся обособлением тех или иных зачатков проспективные способности клеток этих зачатков ограничиваются, и при полном удалении какого-либо зачатка зародыш развивается с соответствующим дефектом. Таким образом обособившиеся зачатки не могут замещать друг друга, их судьба строго предопределена даже уже тогда, когда видимой дифференцировки нельзя еще обнаружить.
Детерминация ведет через дифференцировку клеток и зачатков к развитию строго определенной структуры организма.
В некоторых случаях и число клеточных делений данного зачатка точно предопределено, и соответствующий орган взрослого животного состоит тогда из определенного числа определенным образом размещенных клеток.
Само собой разумеется, что действительно учесть число клеток мы можем только у микроскопически малых животных. У других животных огромное число клеток, составляющих любой орган, делает их подсчет совершенно невозможным, и в этом случае можно руководствоваться лишь косвенными указаниями на такую точную определенность структуры данного органа.
Из небольших животных поразительно правильным расположением клеток различных органов отличаются известные тихоходки. У обыкновенной тихоходки — Macrobiotus Hufelandi — глотка, например, состоит всего из 30 клеток, расположенных определенным образом: 6 эпителиальных клеток располагаются по 3 в начале и в конце глотки; между ними располагаются мускульные клетки в виде 4 поперечных серий по б клеток в каждой. Покровный эпителий спинной части тела составляется из 4 рядов по 19 эпителиальных клеток. На границе между желудком и задней кишкой находится группа в 3 железистых клетки. Подобная определенность состава наблюдается и в других органах и в частности в нервной системе.
То же самое наблюдается и у коловраток и в особенности хорошо изучено у прозрачной Hydatina senta (рис. 12), Из многочисленных подсчетов выяснилась точная определенность клеточного строения различнейших ее органов, и это касается не только числа {22}
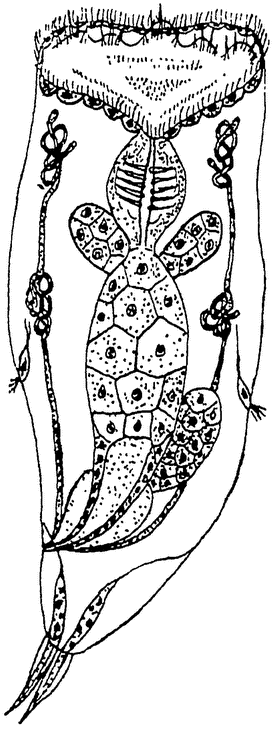 |
Рис. 12. Самка коловратки (Hydatina senta) из Коршельта. Впереди виден коловращательный аппарат, за ним следуют передняя кишка с жевательным аппаратом, желудок с железами и задняя кишка; по бокам выделительные каналы, впадающие в конечный пузырь; справа от желудка — яичник; позади — аппарат для прикрепления. |
Головной |
мозг |
183 |
клетки |
|
Ganglion |
pedale |
23 |
„ |
|
„ |
vesicale |
med. |
2 |
„ |
„ |
„ |
lat. |
9 |
„ |
„ |
ovariale |
post. |
4 |
„ |
2 |
„ |
ant. |
2 |
„ |
„ |
genus |
2 |
„ |
|
„ |
anterius |
8 |
„ |
|
Nervus |
sensualis |
lat. |
2 |
„ |
„ |
„ |
dors. |
8 |
„ |
„ |
lateralis |
inf. |
2 |
„ |
„ |
commissuralis. |
1 |
„ |
|
Всего |
246 |
клеток |
||
Общее число клеток всего животного определено из подсчета ядер всех органов в сумме 959.
И у других коловраток тело слагается, повидимому, также из вполне определенного числа клеток, и то же самое касается и многих других животных. Определенный клеточный состав установлен и для колючеголовых червей и притом сходный даже для различных видов одного рода (Eorhynchus). У некоторых круглых червей установлено постоянство клеточного состава, по крайней мере для некоторых органов, а именно органов чувств, нервной системы и мускулатуры. В других системах органов большое число клеток делает подсчет уже невозможным. Точным подсчетом удалось установить строгое постоянство клеточного состава нервной системы даже таких крупных червей, как аскариды; постоянным оказывается не только число, но и положение, форма и связи нервных клеток. Постоянным является и состав нервной системы пиявок, а по всей вероятности и вообще огромного большинства животных. {23}
У более высоко организованных и крупных животных, каковы моллюски и позвоночные, подсчет даже сравнительно больших нервных клеток совершенно невозможен. Среди большого числа обычных нервных клеток встречаются, однако, особые гигантские нервные клетки. Число последних всегда невелико и точно фиксировано, так же, как их величина, положение, форма, а также и характер ветвлений их отростков и связи их с другими частями нервной системы. Если эти крупные элементы столь постоянны, то, очевидно, не менее постоянны и другие элементы нервной системы высших животных.
Несомненно и органы чувств отличаются постоянством своего клеточного состава. В органе слуха у Triton cristatus в macula neglecta около 107 чувствующих клеток, а у Triton vulgaris — около 83, в papilla basilaris у первого вида тритонов — около 22, а у второго вида — около 11 (G. Рröbsting, 23). Яснее всего это видно, однако, в сложных глазах насекомых, каждый глазной конус которых состоит из определенной системы небольшого числа клеток (например 4 клетки стекловидного тела, 7 клеток ретинулы). Число конусов подсчитать, однако, уже труднее, хотя и не невозможно. Для обыкновенного майского жука оно определяется 5475.
Это число оказывается сравнительно ничтожным, если мы попробуем хотя бы приблизительно учесть число клеток ретины позвоночного. У человека число палочек ретины определяется приблизительно в 250 000 на 1 квадратный миллиметр. Таких величин мы подсчитать не можем, но постоянство клеточного состава, по крайней мере таких органов, как нервная система у мелких животных, дает нам право вывести заключение, что и у высших животных не только общая организация, но и тончайшая структура и клеточный состав этих органов также строго нормированы. Эта строгая определенность только и может быть дана заранее, она должна быть в той или иной форме предопределена, детерминирована интимной структурой яйца. Таким образом, очевидно, строение яйца и соответственная детерминация зачатков ведут к развитию строго определенной структуры организма.
Если организация животного определяется в конце концов внутренними причинами, заложенными в самом, яйце и проявляющимися в дифференцировке отдельных частей яйца или продуктов его деления, то средством для сохранения уже осуществленной организации является регуляция. Под регуляцией мы разумеем способность {24} организма к восстановлению типичного целого при всяких нарушениях нормального соотношения частей.
В большей или меньшей степени регуляторная способность присуща всем организмам и составляет одну из наиболее характерных их особенностей. Так как регуляция осуществляется обычно путем передифференцировки или во всяком случае путем возобновления клеточных делений, то понятно, что она совершается гораздо легче в тех случаях, когда организм мало дифференцирован. В связи с этим стоит правило, что регуляторная способность в общем выше у низко организованных животных. Этим же объясняется и то обстоятельство, что на ранних стадиях развития регуляция формы, много совершеннее, чем на более поздних. Полная и неограниченная регуляция возможна вообще только на самых ранних стадиях деления яйцеклетки (на стадиях дробления) и то в случаях недетерминированного развития, когда яйцо недифференцировано. Примером такой совершенной регуляции могут служить яйца низших кишечнополостных животных, к которым относятся и описанная гидра, и морские полипы, и медузы. При развитии животного из яйца продукты деления последнего могут быть как угодно разделены и перемещены и все же они восстанавливают нормальный организм. Очевидно значение продуктов деления здесь не предопределено их составом, а определяется в зависимости от их взаимоотношения.
Но и на тех стадиях, когда имеется уже известная дифференцировка, и даже у взрослого животного регуляция может быть весьма совершенной. Если у взрослого полипа отрезать любую часть тела, то она свободно восстановится за счет оставшейся части путем более или менее ясной передифференцировки. Гидру можно разрезать на. много частей, и каждая часть восстановится в целую гидру при одном только условии, чтобы в этой части были представлены оба слоя ее тела — эктодерма и энтодерма. Последние друг друга замещать не могут, и таким образом возможность передифференцировки ограничена известными пределами. Такая же высокая регуляторная способность характерна, например, и для пресноводных плоских червей — планарий.
Мы уже указали, что регуляторная способность на ранних стадиях развития в общем выше, чем на более поздних. Известны, впрочем, и исключения: некоторые животные на стадии личинки обладают менее совершенной регуляторной способностью, чем во взрослом состоянии (например личинка Phoronis — Actinotrocha регенерирует значительно медленнее, чем взрослое животное), {25} но в этом случае можно специально личиночные органы рассматривать как уже изношенные, более старые, чем молодое животное, развивающееся из индифферентных ее зачатков. Кажущимся исключением являются еще яйца многих животных с детерминированным развитием, как, например, яйца кольчатых червей или асцидий. В то время как детерминированное развитие круглых червей и пиявок находится в полной гармонии с низкой регуляторной способностью этих животных, у кольчатых червей и асцидий мы находим иное. В этом случае удаление отдельных продуктов деления яйца (бластомеров) приводит к образованию зародыша с известными дефектами. Органы, зачатки которых локализированы в удаленных бластомерах, не могут быть восстановлены. Однако взрослые кольчатые черви и асцидий обладают поразительно высоко развитой регуляторной способностью, и даже из незначительной части их тела легко восстанавливается нормальный организм. В данном случае нужно, однако, принять во внимание, что при удалении определенного бластомера мы совершенно удаляем зачаток того или иного листка или системы органов, между тем как во втором случае оставшаяся часть тела содержит все листки и части всех систем органов. Недостающие части развиваются за счет оставшихся частей, в общем в пределах данного зародышевого листка, данной системы органов и даже данной ткани (т.-е. мышцы — из остатков мускульной ткани, нервная система из уцелевших частей нервной ткани и т. д.). Хотя исключения из этого правила и известны, но они не многочисленны и, повидимому, объясняются существованием особых резервных индифферентных тканей.
Таким образом регуляция есть средство для сохранения сложившейся уже организации животного, и является в высшей степени ценным дополнением к детерминации как способу передачи и осуществления этой организации.
Детерминация и строгое фиксирование определенной организации, как оно в особенности выражается в ограничении числа клеток, нормировании их формы, расположения и связей, ведет к изнашиванию отдельных органов, клетки которых не имеют себе заместителей.
Регуляция, наоборот, связана обычно с существованием резервных клеток, которые могут быть выдвинуты взамен выбывших из строя. Если детерминация и связанная с нею дифференциация ведут к старости, то регуляция есть одно из средств борьбы организма, со старостью. {26}
Изнашивание организма выражается в проявлении целого ряда функциональных расстройств, характеризующих старческий период жизни. Мы уже видели, что и у простейших несмотря на их потенциальное бессмертие наблюдаются явления старческого вырождения. Здесь старость не связана со смертью, но устраняется периодическим обновлением организации. В чем же заключаются явления старости?
У простейших явления вырождения культур наблюдали уже Мопа, Гертвиг и Бючли, но в особенности детально они были изучены Калкинсом. Характерным для стареющих культур является в особенности общее падение жизнедеятельности, выражающееся в ослаблении обмена, в падении интенсивности размножения и общей вялости; размеры тела уменьшаются, протоплазма становится плотной, зернистой и потому мало прозрачной; различные клеточные органы дегенерируют; большое ядро, сохраняя свою прежнюю величину, кажется относительно более крупным; малое (генеративное) ядро явно гипертрофируется. Так как руководящая роль ядра во всех жизненных процессах обнаруживается вполне ясно, то все авторы обращают особое внимание на изменения ядерного аппарата и в нем ищут причину всего явления. Уже Мопа говорит о старческом вырождении ядер, а Гертвиг видит причину наблюдаемых явлений в нарушенном соотношении объемов ядра и протоплазмы. Калкинс полагает, что причиной наблюдаемой депрессии является истощение известного запаса физиологической энергии, наступающее особенно быстро при однообразных условиях питания. При изменении питания возможно, однако, новое возбуждение жизненной энергии— омоложение организма. Такое омоложение возможно по Калкинсу лишь в известных пределах. Наконец наступает полное истощение потенциальной энергии зародышевой плазмы, что выражается в особенности в утрате способности размножения, и тогда животное умирает от старости.
В настоящее время все эти явления рисуются в несколько ином свете. Мы знаем из опытов Вудрёф и Эрдманн, что период депрессий кончается внутренней перестройкой ядерного аппарата, и таким образом центр тяжести не в изменении питания, которое само по себе не является омолаживающим фактором, а в глубокой реорганизации. Кроме того мы знаем, что смерть от старости при достаточно тщательном ведении культур совершенно {27} исключается. Дегенерация культур у прежних исследователей объясняется, как теперь думают, вредным влиянием накопления известного количества продуктов обмена. Перестройка ядерного аппарата ведет к полному обновлению организации, к полному омоложению. Однако факт периодического возвращения состояния депрессии, т.-е. состаривания культур даже при идеальных условиях их содержания, несомненен, и причина этого лежит очевидно в изнашивании ядерного аппарата.
У сложных животных явления старости в большинстве случаев очень мало изучены. Вполне ясно они бросаются в глаза у млекопитающих и в частности у человека. Как известно, обычными спутниками старости являются поседение волос, утеря эластичности соединительной ткани, в частности в коже (благодаря чему появляются обильные морщины), в кровеносных сосудах (явления артериосклероза), в легких. Мускулатура дегенерирует, и следствием этого является все прогрессирующая мышечная слабость. Развивается дальнозоркость. Масса костей уменьшается вследствие их постепенного рассасывания, мозговые полости увеличиваются, компактное вещество истончается, губчатое становится более крупноячеистым. Кости приобретают чрезвычайную хрупкость. Жизненно важнейшие органы, как мозг, сердце, печень, почки, селезенка, постепенно атрофируются, как это видно уже из простого уменьшения их веса, и в особенности это касается также желез внутренней секреции (половой железы, щитовидной, гипофизы, околопочечников). Благодаря всему этому, а в особенности в связи с атрофией нервной системы и с потерей эластичности стенок кровеносных сосудов, кровообращение, а следовательно и питание различных органов, нарушается, функциональная деятельность всего организма расстраивается, и общий жизненный тонус явно падает.
Все эти изменения являются весьма глубокими и касаются собственно всего организма в целом. Стареют органы, ткани и клетки. Хотя особенно ясно старческая дегенерация бросается в глаза в наиболее дифференцированных органах и тканях, как нервная система, органы чувств и мускулатура, но в самом деле и мало дифференцированные и даже регенеративные ткани не избегают общей участи.
Имеется много наблюдений, указывающих, что старый организм вообще реагирует иначе и находится в ином положении, чем молодой, и притом, повидимому, во всех отношениях. Гартлауб наблюдал изменения в способе образования медуз в стареющей колонии гидроидных полипов (Syncoryne), которая каждый год отмирала {28} и регенерировала вновь из остатков ценосарка. Постепенно эта колония совершенно утратила способность образовывать медузы. Чайльд показал, что молодые ресничные черви — планарии — быстрее приспособляются к жизни в слабых растворах алкоголя, чем старые, и регенерация у старых идет медленнее, чем у молодых.
Лишь у немногих животных старческие изменения прослежены более детально. Так, например, Гармс (12) описывает старческую дегенерацию нервной системы у червя Hydroides pectinata. В связи с дегенерацией нервной системы здесь стоит и дегенерация других органов, как кровеносных сосудов и кишечника. У коловраток старческие изменения описаны Шпеманном (23).
Очень интересны исследования Коршельта (20) и. Вальтера (21) над продолжительностью жизни и старением у низших ракообразных (Copepoda и Ostracoda). У циклопов прежде всего бросаются в глаза глубокие изменения в нервных клетках мозга, ведущие затем к постепенному их разрушению. Дегенерирует, между прочим, и лопасть мозга, снабжающая первую пару антенн. В связи с этим дегенеративные изменения распространяются и на последние. Начиная с дистальных частей, в антеннах развивается значительная пигментация, и происходит распад тканей, в особенности мускулатуры; значительно изменяются и хитиновые части, так что внешний вид их резко меняется. В кишечнике старческие изменения начинаются также довольно рано и обнаруживаются в накоплении в клетках его эпителия все больших количеств конкрементов мочекислых солей, что указывает на несовершенное удаление продуктов обмена у стареющего животного. Затем наступает и настоящая дегенерация эпителия кишечника. Все эти изменения сопровождаются постепенным угасанием всех жизненных функций. Так как антенны служат у циклопов органами движения, то их дегенерация ведет к потере подвижности. Животное в конце концов падает на дно и постепенно умирает. Старческая дегенерация описывалась и у насекомых (Dyliscus marginalis, Вlunсk, 23)*).
Однако все же наиболее глубоко проанализированы явления старческой дегенерации органов и тканей у человека. И здесь, повидимому, впереди идет нервная система. Дегенерация мозга выражается даже в уменьшении его объема и веса. Извилины головного мозга становятся уже, а борозды между ними раздвигаются шире. Полости желудочков увеличиваются. Изменяются мозговые оболочки и кровеносные сосуды. Нервные клетки дегенерируют, и работоспособность всего нервного аппарата значительно понижается. {29}
Одновременно наблюдаются глубокие изменения в кровеносной системе. Стенки кровеносных сосудов теряют свою эластичность, и в них происходят отложения извести — явления, известные под названием артериосклероза. Хотя подобные же изменения, сопровождающиеся также и отложением извести, наблюдаются и в других тканях и органах, но здесь они особенно ясно нарушают функцию одной из важнейших систем органов. Потеря эластичности сосудов ведет к затруднению кровоснабжения отдельных органов и вызывает компенсаторную гипертрофию сердца. Усиленная работа последнего, очевидно, ускоряет ход старческих изменений в самом сердце. Хрупкие стенки сосудов под давлением крови легко разрываются, и наступают кровоизлияния в те или иные, иногда жизненно необходимые, органы, что уже сразу может оказаться смертельным. Старческие изменения в самом сердце выражаются в постепенной дегенерации сердечной мышцы и других не менее важных изменениях, которые нередко являются прямой или косвенной причиной наступающей смерти.
Однако сердце позвоночных животных, в частности человека, вообще говоря, оказывается чрезвычайно стойким, как это показывают многочисленные эксперименты с переживанием частей вне организма. Устанавливая питание сердечной мышцы при помощи питательных жидкостей, проводимых по, собственным сосудам сердца, удавалось оживить сердце человека, вынутое из трупа, спустя сутки, а у животных даже через 3—4 суток. Поэтому старческое ослабление сердечной деятельности в настоящее время ставят в связь с дегенеративными изменениями в нервной системе. Последняя играет таким образом руководящую роль, и смерть от старости зависит прежде всего от атрофии центральной нервной системы (Милльманн, Рибберт).
Старческие изменения в нервной системе выражаются гистологически в дегенерации отдельных нервных клеток. Но что же вызывает эту дегенерацию? Мы уже неоднократно говорили об „изнашивании” дифференцированных элементов, но ни разу еще не пытались проникнуть в глубь этого явления, чтобы уяснить себе характер и неизбежность этого изнашивания.
У циклопов Вальтер (21) проследил ход старческих изменений в нервных клетках их центральной нервной системы. По сравнению с молодыми животными изменения в строении нервных клеток довольно значительны: прежде всего замечается скопление каких-то сильно красящихся зернистых масс в окружности ядра, {30} но затем происходят дегенеративные изменения и в самих ядрах; хроматин, первоначально рассеянный в виде отдельных зернышек по ядерной сети, собирается в более крупные комья и, наконец, образует одну сплошную хроматиновую массу в центре или на периферии ядра. Наконец исчезает хроматиновая масса, растворяется оболочка ядра, и как ядро, так и плазма нервной клетки распадаются, частью при явлениях вакуолизации.
Подобную дегенерацию нервных клеток наблюдал Годж (95) и у пчел, причем по его расчету у старых пчел сохраняется лишь около 1/3 части нервных клеток, имеющихся у молодой пчелы.
 |
Рис 13. Разрез через червячок мозжечка 2-летней собаки по Гармсу из Коршельта. gz — нервные клетки (Пуркинье). |
И эти тонкие изменения в строении нервных клеток изучены лучше всего у млекопитающих, в частности у человека.
Гармс детально исследовал гистологическое строение мозга у собак различного возраста. Оказывается, с возрастом в протоплазме нервных клеток накапливаются отложения пигмента, особенно в пирамидных клетках большого мозга, а затем идет и дегенерация нервных клеток, которая была особенно заметна в Пуркиньевых клетках мозжечка (рис. 13, 14). Нервные клетки уменьшаются в раз. мерах; хроматин сбивается в комья, и, наконец, наблюдается настоящий распад клеток (рис. 15). Тело клетки переполняется жировыми капельками, нервные фибрилли и тельца Ниссля исчезают, а также исчезают и клеточные отростки; контуры ядра теряются, и вся клетка превращается в бесформенную массу, которая затем поглощается фагоцитами. Фагоциты и нормально имеются в нервной {31}
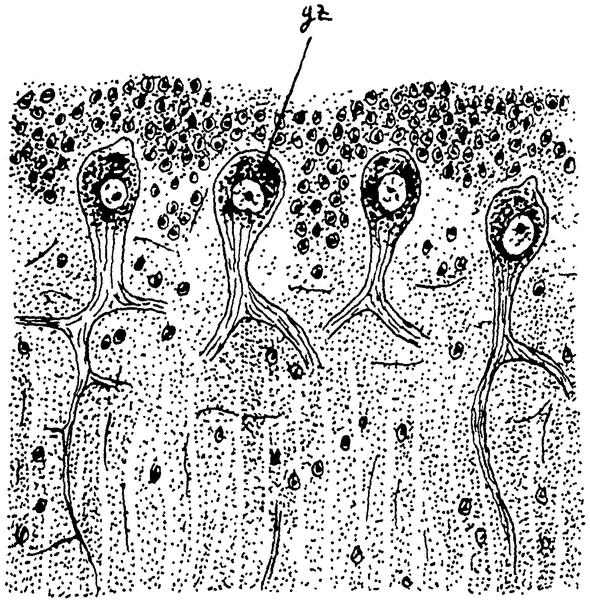 |
Рис. 14. Такой же разрез через червячок мозжечка 14-летней собаки по Гармсу из Коршельта. Нервные клетки (gz) наполнены пигментом. |
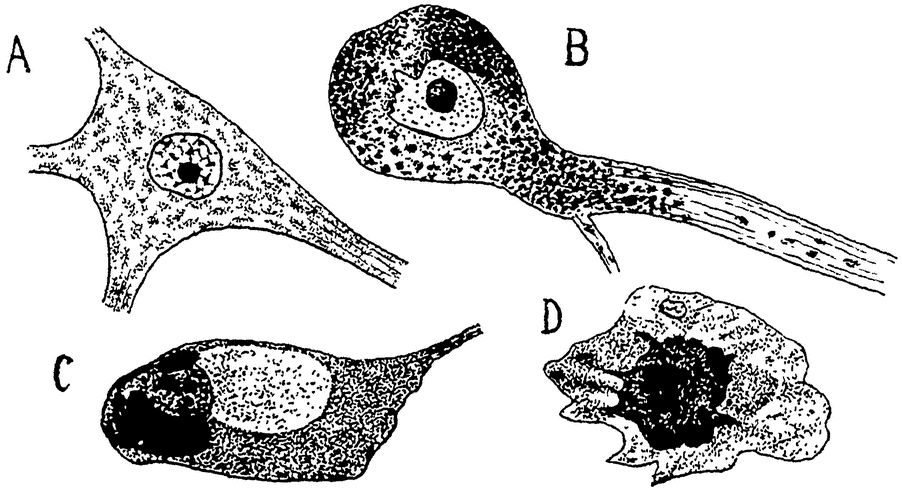 |
Рис. 15. А — нервная клетка из брюшного рога спинного мозга (шейной области) 2-летней собаки, с ясными тельцами Ниссля. В — нервная клетка из пограничной области продолговатого и спинного мозга 12—13 летней собаки. Клетка переполнена липоидными зернами. С, D — нервные клетки шейной области спинного мозга 17-летней собаки в состоянии дегенерации; слева масса липоидного пигмента, справа распадающееся ядро. По Гармсу из Коршельта. |
| {32} |
В нервных клетках человека ход старческих изменений прослежен в особенности Милльманном. Уже в нервных клетках детского мозга появляются включения в виде мелких липоидных пигментированных зернышек, равномерно рассеянных по всему телу клетки. С возрастом число таких зернышек возрастает, они становятся крупнее и образуют местные скопления в виде более или менее значительных конгломератов. В нервных клетках стариков наблюдались очень значительные массы „старческого пигмента”.
То же самое явление наблюдалось и у других млекопитающих и у птиц. Гармс, как уже говорилось, детально проследил накопление липоидного пигмента в нервных клетках собаки. У старых собак последние бывают совершенно набиты пигментными зернами.
Это накопление „старческого пигмента” является, понятно, лишь ясно видимым проявлением более глубоких процессов. По всей вероятности, в результате недостаточного удаления продуктов обмена, они накапливаются в том или ином виде (пигмент) в теле клетки и оказывают, как это признается вообще для продуктов обмена, вредное влияние на жизнедеятельность клетки. В результате клетки испытывают нечто вроде очень постепенного и медленного жирового перерождения. Клетки сморщиваются, теряют свои отростки, вакуолизируются, ядра их, как мы уже видели, явно разрушаются. Наконец происходит полный распад и уничтожение остатков при помощи фагоцитов.
Большое значение фагоцитов в разрушении нервной системы подметил уже И. Мечников, однако он, пожалуй, переоценил его, приписывая фагоцитам слишком активную роль в уничтожении ослабленных нервных элементов. Очевидно, как и в других органах, фагоциты играют лишь роль ассенизаторов, уничтожающих только погибшее, уже безжизненное.
Таким образом в явлениях старческих изменений организма руководящую роль играет нервная система. В этой последней, по А. С. Догелю, особенно большое значение имеет, однако, не центральная, а симпатическая нервная система, которая во всех явлениях дегенерации идет впереди. В симпатической нервной системе человека, как и в центральной, клеточные деления прекращаются уже около времени рождения. Так как симпатическая нервная система ведает прямо или косвенно отправлениями всех органов тела, то жизненные функции организма регулируются прежде всего деятельностью этой системы. Во многих частях организма работа симпатической {33} нервное системы не прерывается периодами отдыха (как это имеет место в центральной нервной системе во время сна) и является совершенно непрерывной и бессменной. Неудивительно поэтому, что дегенерация клеток симпатической нервной системы начинается еще раньше, чем в мозгу. Даже у зародышей встречаются уже отдельные перерождающиеся клетки. После рождения с возрастом все более увеличивается число гибнущих клеток, остатки которых поглощаются фагоцитами. К старости дегенерация значительно подвигается, и перерождаются сразу целые группы клеток. На месте исчезнувших клеток разрастается соединительная ткань.
В результате этих процессов старческой дегенерации наблюдается весьма значительное уменьшение числа функционирующих нервных клеток. По подсчету Гармса, у 14-летней собаки число Пуркиньевых клеток мозжечка равняется приблизительно 2/3, а у 17-летней падает — до 1/3 общего их числа, имеющегося у молодой 2-летней собаки. Однако и эти сохранившиеся еще клетки уже значительно изменены и таким образом функциональная деятельность нервной системы, понятно, является глубоко подорванной. В связи с падением деятельности симпатической нервной системы наблюдаются резкие нарушения кровообращения, а следовательно и питания отдельных органов, нарушение деятельности желез внутренней секреции и т. д. К этому нужно прибавить, что старческая дегенерация клеточных элементов наблюдается и в других органах, в том числе и в железах внутренней секреции, что опять-таки резко отзывается на всей жизнедеятельности организма.
Дегенеративные изменения в клетках представляют собой, повидимому, результат ненормального обмена. Продолжительная деятельность клеток ведет к накоплению продуктов обмена, оказывающих вредное воздействие на живое вещество1); ясно видимым выражением этого процесса постепенного самоотравления является прежде всего, как уже указывалось, накопление старческого пигмента. Эти отложения пигмента наблюдаются с возрастом в различных тканях, но особенно рано, а позже в значительных массах этот пигмент накапливается, как уже сказано, в нервных клетках (особенно {34} в симпатических и спинномозговых узлах), затем в сердечной мышце, в поперечнополосатой мускулатуре (особенно органов дыхания) и в гладкой мускулатуре (особенно кровеносных сосудов и кишечника).
Таким образом вполне ясно старческие изменения наблюдаются в наиболее дифференцированных тканях организма. Однако они несомненно касаются и всех вообще тканей. Известно, что регенерационная способность с возрастом неуклонно падает. Так как заживление ран происходит при активной деятельности мало дифференцированных частей соединительной ткани и эпителия, то, следовательно, и эти элементы теряют свою жизнеспособность. То же самое
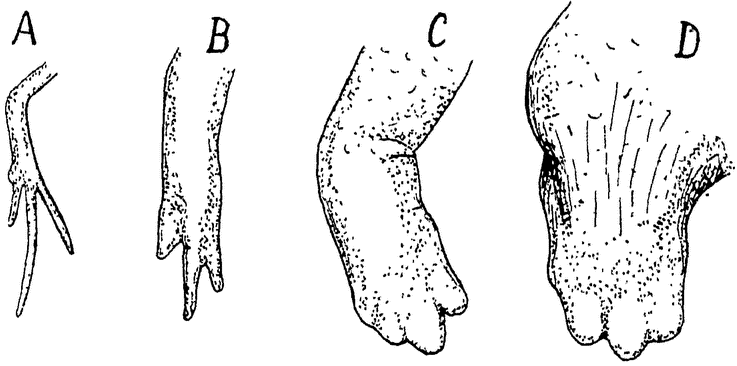 |
Рис. 16. Зависимость регенерации передней конечности тригона (Triton cristatus) or возраста. А — регенерация у молодой личинки (11 мм до заднепроходного отверстия); В — регенерация у более поздней личинки (25 мм длины); С — регенерация у большой личинки (40 мм длины); D — регенерация у взрослого животного. На всех рисунках изображена приблизительно одинаковая стадия развития (В — немного моложе других) и, при одном увеличении (6 раз). |
показывают и эксперименты с тканевыми культурами. Хотя клетки соединительной ткани старых животных и не теряют вполне своей способности к размножению, но интенсивность этого размножения и рост культуры всегда гораздо ниже, чем у молодых животных. Таким образом и мало дифференцированные, регенеративные ткани также подвержены старческим изменениям. Интересно, что падение регенерационной способности проявляется уже очень рано, во время эмбрионального развития, и является, пожалуй, наилучшим выражением существования возрастных изменений живого вещества, связанных с наличностью жизненных процессов организмов (рис. 16).
Возрастные изменения живого вещества несомненно покоятся на физико-химических его преобразованиях. По исследованиям Ружички (Ruzicka, V. 1922) вещество живых организмов от начала {35} развития до смерти непрерывно сгущается („Protoplasmahysteresis”). Эта прогрессивная конденсация представляет собой важнейшее старческое изменение, так как она является уже физическим препятствием для процессов обмена, а ограничение обмена влечет за собой, как мы видели, и все прочие явления старческой дегенерации. Очень важно то обстоятельство, что степень конденсации живого вещества оказывается измеримой, и таким образом в наших руках имеется уже средство для определения истинного возраста живого вещества данного организма или отдельного его органа.
У простейших устранение явлений старости достигается периодическим обновлением внутренней организации и в частности ядерного аппарата, связанным с процессом размножения.
У сложных животных такое обновление организма недостижимо, так как восстановление одних функций живого вещества без одновременного возобновления других, очевидно, невозможно. Полное омоложение всех клеток сложного организма путем перестройки их ядерного аппарата привело бы, конечно, к полному возрождению и прежде всего к восстановлению интенсивного обмена веществ, но вместе с тем преобладание ассимиляции над диссимиляцией, характерное для молодого организма, неизбежно связано с нарастанием живого вещества, размножением клеток и т. д. Этому последнему имеются, однако, естественные препятствия — организмы с неограниченным ростом совершенно немыслимы в реальных условиях существования.
Таким образом неизбежность установления предела роста ограничивает и жизнь известными пределами. Постепенное приближение к этому пределу и сопровождается обыкновенно явлениями старости.
Борьба организма со старостью возможна, казалось бы, еще путем постепенного замещения старых, уже, изношенных, клеток более молодыми, которые сохранялись бы в виде особого резерва мало дифференцированных и не функционирующих тканей. Такие резервные ткани действительно имеются в организме у многих животных и служат для смены быстро изнашивающихся частей организма.
Совершенно ясно наблюдается такая непрерывная смена, например, в коже высших позвоночных (рис. 17). Эпидермис позвоночных, как известно, является в виде многослойного эпителия, верхние {36} слои которого состоят из плоских клеток, постепенно изнашивающихся и отмирающих. Нижний слой эпидермиса состоит из цилиндрических клеток, сохраняющих эмбриональный характер. Эти клетки размножаются путем деления, и продукты этого деления перемещаются кнаружи, подвигаясь на смену отмирающим клеткам. Особенно ясен весь этот процесс у наземных позвоночных, у которых клетки наружных слоев эпидермиса испытывают особое роговое перерождение, превращаясь в мертвые плоские роговые чешуйки, образующие прочные связные пласты, которые защищают тело животного от всяких вредных воздействий внешней среды. Этот „роговой” слой эпидермиса может достигать значительной мощности, особенно
 |
Рис. 17. Разрез через кожу млекопитающего: со — кожа собственно; b — основной слой эпидермиса, sc — роговой; sM — Мальпигиев слой. |
Роговой покров кожи не является постоянным, он сменяется или путем непрерывного слущивания верхних слоев, как, например, у млекопитающих, или путем отделения больших лоскутов или даже всего наружного слоя рогового покрова в виде цельного образования, как при линьке ящериц и особенно змей. Более сложные роговые образования, как перья и волосы, также периодически сбрасываются и заменяются новыми.
Лежащий под роговым слоем живой и богатый мало измененной протоплазмой „Мальпигиев” слой непрерывно возобновляется путем размножения клеток нижнего его слоя. Этот нижний слой клеток имеет характер индиферентной эмбриональной ткани, и его деятельность является непрерывной, хотя в старости и эта деятельность постепенно угасает (например смена и рост волос).
Подобные же процессы происходят и в других органах, хотя они не всегда так ясно обнаруживаются. Так, например, идет смена клеток во многих железах. В особенности постоянное разрушение клеток при явлениях жирового перерождения и возобновление их из нижних слоев эпителия наблюдаются в сальных железах кожи {37} млекопитающих. В млечной железе наблюдаются периодическое изнашивание и редукция всей железы и затем новое ее восстановление. В эпителиальной выстилке кишечника также происходят постоянное изнашивание клеток и замена их другими, более молодыми.
Наконец, форменные элементы крови, в особенности красные кровяные шарики млекопитающих, имеют весьма преходящее значение. Эти клетки, как известно, в развитом состоянии теряют свои ядра, являясь весьма специализированными образованиями. В связи с этим они не только теряют способность размножения, но и вообще их существование ограничено ничтожным сроком в 2—3 недели. По мере гибели этих элементов они заменяются новыми, образующимися путем размножения настоящих клеток с ядрами в так называемых кровотворящих органах, как селезенка и костный мозг.
Возможно, что такое постепенное замещение изнашивающихся клеток другими играет весьма значительную роль и в других органах; однако в некоторых органах, и именно в наиболее высоко дифференцированных, как, например, в нервной системе и в существенных частях органов чувств у высших животных, эта смена несомненно во взрослом состоянии не наступает. Отсутствие смены элементов в высоко дифференцированных органах объясняется, очевидно, точной координацией функций отдельных частей и невозможностью их смены без временного нарушения жизненных функций всего организма.
Очень большое значение имеют резервные ткани в тех случаях, когда индивидуальная жизнь разделяется на определенные периоды, характеризующиеся различными соотношениями организма с внешней средой. В этом случае каждый период имеет свою точно определенную организацию, и переход от одного периода к другому может быть связан со значительными преобразованиями — уничтожением старых органов и развитием новых за счет резервных тканей.
Наиболее ярким примером этого явления могут служить насекомые, жизнь которых более или менее резко разделяется на 2 главных периода — личиночный и имагинальный (взрослая форма). Относительно очень длинный период личиночной жизни характеризуется совершенно определенной и вполне законченной организацией. Организм, приспособленный к определенным условиям существования, лишенный лишь развитых половых органов — личиночная форма, — обладает ограниченным ростом. По достижении определенных размеров рост прекращается, и личинка переходит в стадию {38}
 |
Рис. 19. Кишечник куколки мухи с имагинальными зачатками (обозначены черным) — слюнных желез (s), передней кишки (v), эпителия (ie) и мускулатуры средней кишки (lm), задней кишки (е), кожи брюшка (a). По Ковалевскому из Коршельта и Гейдера. |
Если принять во внимание значительную продолжительность жизни личинки (годы и иногда даже десятки лет), при которой
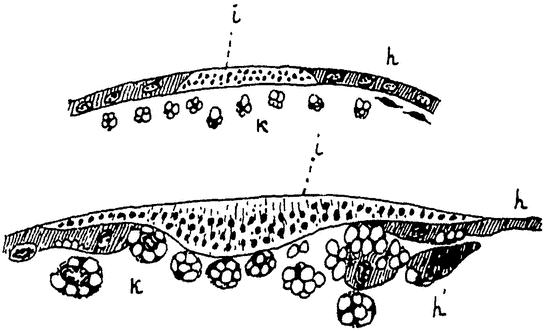 |
Рис. 18. Имагидальный диск в коже личинки мухи. Диск (i) разрастается и вытесняет старую личиночную кожу (гиподерму h, отмечена штриховкой), которая разрушается фагоцитами (k). По Ковалевскому из Коршельта и Гейдера. |
все ткани могли бы уже совершенно износиться, то можно понять, что не будь этой замены, этого обновления организации, готовое насекомое собственно рождалось бы на свет уже совершенно старым и изношенным. Впрочем, оно до известной степени так и есть. Дело в том, что не все органы испытывают во время метаморфоза полное обновление. Даже при наиболее совершенном метаморфозе сохраняются сердце и нервная система, хотя они и испытывают {39} известные преобразования. Таким образом, если взрослое насекомое начинает свою жизнь в общем с обновленной организацией, то все же оно не сравнимо с молодым моллюском или позвоночным: запас жизненной энергии уже до известной степени израсходован, и в особенности нервная система уже несколько истощена. По всей вероятности, этим и объясняется незначительная продолжительность жизни взрослых насекомых.
И во многих других случаях индивидуальное развитие животных связано с глубоким метаморфозом, при чем окончательная форма строится за счет резервных эмбриональных тканей. Так как органы личинок обнаруживают иногда довольно высокую специализацию, то такое глубокое обновление организма перед началом зрелого периода жизни имеет, разумеется, глубокий смысл. Нельзя, однако, сказать, что мало дифференцированные резервные ткани совершенно избегают состаривания. Хотя они и не несут каких-либо специальных функций, но все же участвуют в общих процессах обмена. Таким образом процесс старения должен до известной степени отзываться и на этих тканях. Наблюдения над регенерацией у животных различного возраста, а также и опыты с культурами тканей вне организма показывают ясно, что и мало дифференцированные ткани старых животных растут гораздо медленнее, чем ткани молодых.
Из всего сказанного ясно, что высоко дифференцированные ткани вообще изнашиваются и стареют быстрее, чем мало дифференцированные и не потому, что их клетки являются менее стойкими, а потому, что они не имеют себе заместителей.
Это быстрое изнашивание тканей постоянного состава выражается в большинстве случаев довольно ясно в дисгармоничном старении отдельных органов. Мы не будем говорить о дисгармоничном старении патологического характера, как, например, преждевременное поседение или преждевременный артериосклероз, что нередко наблюдается. Нас здесь в большей мере интересует, например, довольно постоянное явление раннего изнашивания нервной системы. Нет ли способов борьбы с этим негармоничным старением? При невозможности обновления высоко дифференцированных органов для организма остается еще лишь один путь — точная нормировка запаса энергии в виде специфического живого материала соответственно количеству требуемой работы данного органа, При такой точной нормировке негармоничное старение организма может быть устранено. В этом случае в высоко специализированных органах постоянного состава работа должна быть лишь распределена {40} между относительно большим количеством клеток, чем в органах менее специализированных. Таким образом у тех животных, у которых рано изнашивается нервная система (например у млекопитающих), возможно значительное продление жизни при увеличении общей массы этой нервной системы. Как мы увидим в дальнейшем, такая связь между относительной массой нервной системы и продолжительностью жизни действительно во многих случаях установлена.
Таким образом для высоко организованных животных со значительной дифференцировкой отдельных органов остается лишь один путь борьбы со старостью — точная нормировка запаса живого вещества соответственно работе, потребной в течение всей жизни от данного органа. В этом случае старение отдельных органов будет итти равномерно, и таким образом будет достигнута максимальная продолжительность жизни всего организма. Вместе с тем, однако, ясно, что здесь определенность организации ведет к установлению и определенного нормального срока жизни. Высоко организованные животные характеризуются не только точной определенностью своего строения, но и определенной специфической продолжительностью жизни.
Наши сведения о продолжительности жизни различных животных, к сожалению, крайне недостаточны и ненадежны. Приходится считаться с большими различиями в продолжительности индивидуальной жизни особей даже одного вида и с совершенной невозможностью учесть влияние всех факторов, от которых зависят эти различия. Это сказывается даже при попытках установить нормальную продолжительность жизни человека и обычных домашних животных. Тем более возрастают затруднения при переходе к другим животным, для которых содержание в неволе, при котором только и возможно длительное наблюдение, представляет слишком резкое уклонение от нормальных условий существования, которое не может не отразиться на продолжительности их жизни. Мы знаем, что огромное число животных, особенно водных, совершенно не выдерживают жизни в неволе и гибнут в самый короткий срок. Другие живут дольше, и на наблюдениях над этими животными и основываются наши сведения 6 продолжительности жизни различных форм. В большинстве случаев эти данные имеют ничтожное значение, и {41} жизнь в благоприятных условиях существования могла бы быть продлена иногда в десятки раз. Хотя, с другой стороны, средняя продолжительность жизни в неволе бывает, вероятно, заметно выше, чем в естественных условиях, вследствие охраны животных от истребления.
Из приведенных соображений мы не будем придавать большого значения низким показаниям, в особенности для более нежных организмов. Показания больших сроков жизни дают нам, однако, уже более надежный материал. В новейшее время наиболее достоверные сведения о продолжительности жизни животных собраны у Коршельта (24).
Для губок максимальный срок жизни в аквариуме указан в 4 года (Axinella на Неаполитанской станции). В самом деле для крупных колоний срок наверное гораздо выше.
Из кишечнополостных различные колонии гидроидов жили в аквариуме по несколько месяцев, а иногда и несколько лет. И обыкновенная гидра может достигнуть возраста до 2 лет (Стехе, 11). Медузы, естественно, обладают более короткой жизнью, измеряемой только месяцами. Очень крупные сроки указывают для актиний. Эти последние оказываются во многих случаях необычайно стойкими и прекрасно выдерживают содержание в неволе. В аквариумах Неаполитанской станции различные актинии содержались в течение 15—33 лет, а в литературе имеются указания на еще более значительную продолжительность жизни — в 50 (для Sagartia troglodytes) и 67 лет (для Actinia mesembryanthemum). Можно думать, что такая значительная продолжительность жизни связана отчасти с малой интенсивностью обмена, а отчасти с вероятным обновлением организации за счет резервных тканей.
Для ресничных червей литературные указания дают продолжительность жизни в 1 год как для морских Polyclada, так и для пресноводных Triclada. Однако Коршельт указывает, что в его лаборатории пойманные на свободе планарии живут уже до 13 месяцев. К этому я могу прибавить, что у меня около двух десятков Planaria, пойманные осенью 1921 года совершенно взрослыми (я выбирал самых больших), живут в аквариуме без всякого ухода уже 2 года и не обнаруживают никаких изменений. Их возраст выше 2 лет, и нет ничего невероятного в том, что они проживут и еще столько же. Это является лишней иллюстрацией того, насколько велика еще неполнота наших сведений по этому вопросу.
Коловратки обладают несомненно очень коротким сроком жизни. Для Hydatina senta указывают срок в 13 дней для самки и около {42} 3 дней для самца. Rotifer vulgaris достигает возраста 50 дней. Незначительная продолжительность жизни коловраток связана, очевидно, с весьма незначительными их размерами и с ограничением числа клеток всего их организма определенной нормой.
Дождевые черви достигают возраста свыше 10 лет (Коршельт), а пиявки и растут очень медленно и живут еще дольше — 20 до 27 лет (Коршельт).
Асцидиям приписывают сравнительно незначительную продолжительность жизни — в несколько месяцев.
Наблюдений над продолжительностью жизни у иглокожих, к сожалению, имеется очень мало, но так как рост их очень медленный, и половая зрелость нередко достигается в возрасте нескольких лет, то очевидно, что они живут сравнительно очень долго.
Очень значительная продолжительность жизни установлена с несомненностью для многих моллюсков. Обычные наши легочные моллюски живут по несколько лет. Более крупные морские брюхоногие живут еще дольше. Из двустворчатых моллюсков Anodonta и Unio достигают возраста в 10—20, а может быть, и 30 лет. Устрицы живут до 10 лет, а огромные Tridacna до 60—100 лет. Наконец, обыкновенная речная перловица (Margaritana margaritifera), как это с несомненностью доказано случаями вылавливания меченных экземпляров, в раковинах которых был вырезан год предыдущей поимки, достигают поразительного возраста — в 100, а быть может, даже и 150 лет, хотя это и некрупная форма.
Членистоногие живут, вообще говоря, недолго. Мелкие низшие ракообразные живут лишь по несколько месяцев и во всяком случае, повидимому не более 1–11/2 лет, как это доказано обстоятельными наблюдениями Е. Вальтера и Е. Шрейбера, специально поставленными в лаборатории Е. Коршельта. Наиболее ценным результатом этих наблюдений является то, что они установили существование у циклопов (Cyclops viridis) форм с различной продолжительностью жизни. Зимой и ранней весной встречается исключительно форма с нормальной продолжительностью жизни в 9 месяцев. Весной наравне с такими особями начинают появляться и особи с нормальной продолжительностью жизни в 4–5 месяцев. Летом и до осени последняя форма решительно преобладает, а осенью опять начинает увеличиваться численность первой, долго живущей, формы. Эти наблюдения имеют огромное значение для разбираемого нами вопроса, так как показывают ясно, что продолжительность жизни зависит прежде всего от некоторой внутренней конституции {43} организма, которая может и не проявляться в ясных, легко устанавливаемых признаках организации. Более крупные раки живут, понятно, дольше, и, например, обыкновенный речной рак достигает возраста 20–30 лет.
Общая продолжительность жизни насекомых, быть может, и не особенно низка, но своеобразной их особенностью являются весьма значительный личиночный период и ничтожная продолжительность жизни взрослого животного, падающая иногда до нескольких дней, а иногда даже часов, во время которых происходит только спаривание и откладка яиц. Принятия пищи при такой короткой жизни не происходит вовсе. Однако самки муравьев достигают, по указанию целого ряда авторов, весьма солидного возраста — 5–10 и далее 15 лет. У пчел матка также достигает возраста 5–7 лет, но рабочая пчела живет, повидимому, не дольше 6 месяцев, а трутень еще значительно меньше. Здесь мы опять имеем явную зависимость нормальной продолжительности жизни от некоторой внутренней конституции организма. Американская плодовая муха — Drosophila melanogaster — живет в течение 37 дней (Pearl и Parker, 22)*).
У жуков личиночная жизнь тянется обычно в течение нескольких лет, а иногда даже в течение десятков лет. Взрослый жук, наоборот, живет обычно очень недолго, не более 2–3 месяцев, и только у зимующих форм этот срок соответственно увеличивается. В неволе удавалось, правда, содержать крупных жуков в течение ряда лет, но это возможно исключительно при недопущении спаривания. Нормально же самцы погибают через несколько часов или — самое большее — дней после спаривания, а самки — после откладки яиц. Еще короче обычно жизнь бабочек, у которых иногда даже недоразвиваются органы питания, так что они живут исключительно за счет запасов, отложенных в течение личиночной жизни (сатурнии).
Поразительный контраст между продолжительностью жизни личинки и взрослого животного наблюдается еще у цикад, которые во взрослом состоянии живут не дольше нескольких недель, между тем как личиночный период тянется в течение нескольких лет, (не менее 17 лет у американской Cicada septemdecim).
Гораздо больше надежных данных имеется понятно, для позвоночных животных.
Рыбы достигают иногда весьма значительного возраста. Карпы и щуки достигают, повидимому, иногда возраста 150 и даже 250 лет. Довольно точные данные имеются для сельдей, у которых возраст {44} удобно учитывается по числу годичных колец нарастания кожных чешуи. Эти сравнительно небольшие рыбы живут до 15–20 лет. У камбал возраст также легко учитывается (по годичным кольцам на отолитах и на костях жаберной крышки). Эти также не особенно крупные рыбы достигают весьма солидного 50–70-летнего возраста. Не подлежит сомнению, что крупные сомы и осетры, а также акулы обладают весьма продолжительной жизнью. Однако известны и рыбы с коротким жизненным циклом. Бычок — Latrunculus pellucidus — живет только 1 год, умирая после воспроизведения потомства.
Амфибии, по наблюдениям над их жизнью в неволе, достигают возраста 10–15 лет (тритоны, саламандры и лягушки). Жабам приписывают иногда весьма значительное долголетие (до 40 лет).
Относительно большинства рептилий нет достаточных данных. Некоторые ящерицы жили в неволе около 10 лет. Для обыкновенного ужа имеется указание о жизни в течение свыше 33 лет. Крокодилы живут во всяком случае в течение нескольких десятков лет, а крупные черепахи до 200–300 лет. Последние являются, повидимому, животными с наиболее длительной жизнью.
Сравнительно очень долго живут и птицы. Крупные хищники, совы, вороны, попугаи, а также лебеди, достигают и в неволе возраста свыше 100 лет. Большинство даже мелких птиц живут по нескольку десятков лет.
Если долголетие рептилий является вполне понятным при малой интенсивности обмена и слабой общей их жизнедеятельности, то сравнительно еще большая длительность жизни птиц с этой точки зрения не объяснима: это — животное с чрезвычайно интенсивным обменом и весьма высоким общим жизненным тонусом. Продолжительность жизни определяется, очевидно, всей организацией животного и является для него не менее характерной, чем эта последняя.
Относительно гораздо короче жизнь млекопитающих. Мыши и крысы живут до 3–31/2 лет. Морские свинки живут сравнительно долго — до 8 лет. Кролики — 5–7 лет. Крупные хищники — львы, тигры, леопарды — стареют в неволе очень скоро и живут не более 15 лет, но несомненно, что на свободе они живут значительно дольше. Домашняя кошка живет обычно до 10 лет, хотя иногда достигает и вдвое большего возраста. Собаки ясно стареют в возрасте 10–15 лет, хотя также иногда живут значительно дольше. Все это сравнительно очень небольшие сроки, но относительно {45} еще ниже сроки жизни копытных. Овцы и козы живут 10–15 лет; крупный рогатый скот 20–30 лет. При сравнительно большой величине этих животных это несомненно очень короткая жизнь. Немногим больше живет и лошадь. Только такие колоссы среди животных, как слоны, живут, повидимому, около 150–200 лет.
Для обезьян отсутствуют надежные данные, так как эти животные плохо выносят содержание в неволе, однако они, очевидно, достигают сравнительно более высокого возраста. Человек, наконец, обладает также сравнительно высокой продолжительностью жизни: обычно около 70 лет, но иногда и до 100 (или даже выше).
Из приведенного сопоставления, к сожалению, невозможно составить себе цельной и определенной картины. Несомненно, что некоторые закономерности существуют, но они так сложны, что распутать и выделить их не легко.
Прежде всего довольно ясна зависимость между общими размерами тела и продолжительностью жизни. Понятно, что у крупных животных один только период роста длится нередко дольше, чем вся жизнь более мелких.
Однако известно и немало исключений. Крысы живут 3 года (по Ромейсу); мыши несмотря на меньшие размеры живут, повидимому, уже несколько дольше; сравнительно незначительная по величине белка живет 10–12 лет. Для бобра указывают возраст 20–25 лет, т.-е. такой же, как и для крупного рогатого скота.
Еще более бросаются в глаза различия, если сравнивать между собой формы, еще более далеко отстоящие друг от друга. Жабы при очень незначительных размерах живут до 40 лет, т.-е. не менее долго, чем лошадь или верблюд. Наиболее крупные черепахи живут до 200–300 лет. Эти животные живут, следовательно, гораздо дольше, чем млекопитающие. Это может быть объяснено, если принять во внимание их сравнительно слабую жизнедеятельность. Однако и птицы живут очень долго, несмотря на интенсивный обмен и относительно малые их размеры. Среди рыб мы знаем даже не особенно крупных с продолжительностью жизни до 250 лет и, с другой стороны, упомянутого раньше бычка, живущего 1 год.
Не менее значительные различия наблюдаются и между представителями беспозвоночных. Так, обычные наши пресноводные пластинчатожаберные моллюски живут 20–30 лет, другие 2–4 года или даже меньше, однако не более крупная перловица достигает поразительного возраста в 100–150 лет. {46}
Среди насекомых бросаются в глаза различия в продолжительности жизни между, например, ничтожными по своим размерам муравьями, живущими 10–15 лет и сравнительно крупными бабочками и жуками, живущими всего несколько недель (во взрослом состоянии).
Точно так же и среди низших беспозвоночных наравне с ничтожной продолжительностью жизни большинства из них встречаются поразительно высокие числа в несколько десятков лет, как, например, для некоторых актиний.
Таким образом продолжительность жизни несомненно определяется организацией животного, хотя с общей высотой организации она не связана, и у очень близких форм нередко наблюдаются колоссальные различия. Что эти различия действительно предопределяются различной конституцией организма, видно лучше всего по существованию у некоторых видов животных соответствующего полиморфизма. Так, например, полиморфизм у муравьев и пчел связан и с различной продолжительностью жизни: пчела-матка живет до 7 лет, рабочая пчела — несколько месяцев, а трутень — еще меньше и умирает непосредственно после спаривания. Редуцированные самцы коловраток, Dinophilus, усоногих раков живут, конечно, гораздо меньше, чем их самки. У циклопов мы также видели существование двух форм, ясно различающихся продолжительностью жизни (здесь различия не связаны с половым диморфизмом)*).
В некоторых случаях делались попытки уловить связь между организацией животного и продолжительностью его жизни. Первая попытка такого рода принадлежит И. И. Мечникову, который указывал на „дисгармонии” в организации как на фактор, сокращающий жизнь. И. И. Мечников указывал на существование у человека целого ряда органов, которые при современных условиях его существования утратили всякий смысл. Некоторые из них являются рудиментарными и служат вместе с тем явным источником страданий, как, например, червеобразный отросток слепой кишки или зуб мудрости, но иные могут быть еще и вполне развитыми и даже крупными органами. Таковым является, например, толстая кишка человека. Этот значительный орган был необходим для растительноядных предков человека, но теперь утратил свое пищеварительное значение. Процессы бактериального брожения, протекающие в толстой кишке, сопровождаются образованием вредных продуктов распада и бактериальных ядов, которые, постепенно отравляют организм. Если это отравление и не есть причина смерти вообще, то оно во всяком случае значительно {47} укорачивает нашу жизнь. И. И. Мечников доказывает, что и у других животных, у которых толстая кишка, быть может, и является необходимым органом, она также является фактором, сокращающим жизнь.
Млекопитающие с длинной толстой кишкой, как, например, жвачные и лошадь, живут, как мы видели, сравнительно очень недолго. Летучие мыши, обладающие очень короткой толстой кишкой, живут дольше, чем другие млекопитающие равных размеров. У птиц, продолжительность жизни которых обращает на себя особое внимание, имеется совершенно незначительная толстая кишка. Сравнительно недолго живут лишь страусы, но у них-то мы и находим опять сравнительно сильно развитую толстую кишку. Низшие позвоночные (рыбы, амфибии, рептилии), обладающие лишь незначительной толстой кишкой, живут также сравнительно долго.
Очень интересную закономерность подметил еще Фриденталь, который учитывал отношение веса мозга к весу остального живого вещества. Это отношение является наивысшим у человека, затем у человекообразных обезьян, у слона и у других животных с высоко развитыми умственными способностями. Оказывается, этот наивысший „фактор цефализации” совпадает и с относительно наивысшей продолжительностью жизни. Наиболее умные животные производят наибольшую жизненную работу и живут дольше других.
Все это пока еще лишь первые попытки установить те особенности организации, которые более или менее непосредственно определяют долголетие животного. Несомненно, наиболее интересной попыткой является эта последняя и именно потому, что она, быть может, окажется имеющей более общее значение. Нервная система по существу своему является бессменно работающим регуляторным аппаратом, и в нем, как мы видели, обычно раньше всего обнаруживаются явления старческой атрофии. Если работа, падающая на этот центральный аппарат, будет распределяться между бóльшим числом клеток, т.-е. будет протекать в большей массе живого вещества, то изнашивание нервной системы, разумеется, обнаружится значительно позже; таким образом понятно, что относительно большей массе нервной системы обычно соответствует и бóльшая продолжительность жизни. Весьма вероятно, однако, что значение различных частей нервной системы окажется в этом отношении далеко не одинаковым. Мы видели, что у позвоночных животных для жизненных функций организма особенно большую роль играет симпатическая нервная система. Надо ожидать, что масса именно {48} этого отдела нервной системы имеет решающее значение как фактор, определяющий долголетие, между тем как масса высших центров психической жизни, очевидно, не играет заметной роли, по крайней мере при определении срока наступления физиологической смерти (по биологическим условиям атрофия высших центров, разумеется, может фактически привести к очень быстрой насильственной или голодной смерти).
Всегда ли, однако, увеличение массы нервной системы животного будет приводить к продлению его жизни? Разумеется, нет. Явления старческой атрофии наблюдаются и на других органах, и для них можно было бы повторить все сказанное для нервной системы. Ясно, что увеличение массы нервной системы имеет смысл только до тех пор, пока она оказывается относительно недостаточной, что, повидимому, в самом деле нередко имеет место (ввиду отсутствия резервных клеток). Максимальная продолжительность жизни достижима, очевидно, лишь при установлении точнейшего соотношения между массами функционирующих в течение всей индивидуальной жизни тканей (т.-е. считая и вновь продуцируемые за счет резервных тканей) различных органов. Иными словами, условием максимального долголетия является строгая гармоничность всей организации.
Понятно, что такая физиологическая гармония может и не совпадать с биологическими требованиями, предъявляемыми к организму, и у животных при наличии борьбы за существование, когда естественной смерти сплошь да рядом реально и не существует вовсе, во многих случаях гораздо важнее иметь биологически совершенную организацию („приспособленную” к данным условиям существования). Удовлетворение биологических требований может скорее привести к фактическому1) удлинению жизни в естественной обстановке, чем физиологическая гармония организации. С этим, вероятно, и связано обычно наблюдаемое дисгармоничное старение животных, содержимых в искусственных условиях.
Наконец, вовсе не является доказанным, что долголетие является биологически „полезным” признаком. Во многих случаях наблюдается какое-то странное, как будто насильственное устранение {49} организмов с жизненной арены. Это — смерть физиологическая, „естественная”, но вместе с тем ее нельзя назвать смертью от старости. Особенно странное впечатление производит быстрая смерть большинства насекомых после спаривания. Такая смерть обыкновенно наблюдается у обоих полов, но иногда оказывается связанной лишь с одним определенным полом. Так, у муравьев, матка которых отличается поразительным долголетием, самцы умирают сейчас же после спаривания (у биологически сходных термитов самец живет, однако, в течение нескольких лет и спаривается многократно). У пчел, матка которых живет в течение 5—7 лет, трутни умирают во время свадебного полета, тотчас же после спаривания (в течение нескольких секунд). Общая продолжительность жизни во взрослом состоянии может быть таким образом в высшей степени сокращенной. Наиболее известным и ярким примером такой краткой жизни являются поденки (Ephemeridae), жизнь которых иногда измеряется только часами. После вылупления из куколки поденки вечером поднимаются на воздух, спариваются, самки откладывают яйца, и ночью же они уже умирают. Таким же образом бабочки из семейства Psychidae партеногенетически откладывают яйца непосредственно по вылуплении из куколки и после этого умирают.
В некоторых случаях смерть после воспроизведения потомства является смертью от истощения. Так, например, это можно было бы предположить для некоторых рыб (кэта), откладывающих массу яиц, для самок многих бабочек, мух, прямокрылых, для самок поденок и т. д., но вряд ли это объяснение применимо к самцам тех же животных. В других случаях это, очевидно, смерть от крайнего напряжения, приводящего к параличу нервной системы („Shocktod”, Дофлейна), как, например, смерть трутней пчел связана с напряжением при выпячивании их огромного копулятивного органа; вероятно то же касается и самцов муравьев. Наконец, во многих случаях у насекомых смерть есть результат недостаточного питания или даже голода вследствие того, что ротовые части и органы пищеварения оказываются редуцированными. Взрослые насекомые живут главным образом за счет запасных материалов, накопленных в течение личиночной жизни.
Во всех этих случаях мы, однако, опять возвращаемся к особенностям физиологической конституции организма, как к фактору, определяющему продолжительность жизни. Нам приходится отказаться от предвзятой мысли о „полезности” длительной жизни и признать, что. для каждого вида животных (и даже каждого пола и каждой {50} формы) имеется своя характерная и притом вовсе не максимально возможная продолжительность жизни, определяемая всей его организацией. В некоторых случаях наблюдается поразительно краткая жизнь, явно определяемая особой предустановленной дисгармоничностью физиологической конституции.
Если продолжительность жизни связана с организацией (физиологической конституцией) животного, то вместе с тем ясно, что она, как и последняя, определяется сложным комплексом биологических факторов и, следовательно, может быть рассматриваема как приспособление.
В особенности известна попытка Вейсманна объяснить с этой точки зрения наблюдаемые различия в продолжительности жизни различных животных и в частности, например, птиц и млекопитающих. Ясно, что отдельные особи любого вида животных должны существовать столько времени, сколько нужно для того, чтобы обеспечить существование данного вида воспроизведением достаточного потомства. Если эта норма жизни падет ниже, то вид неизбежно вымрет. Вместе с тем срок жизни обычно не превышает этой нормы, так как и сама смерть, по Вейсманну, есть полезное приспособление, устраняющее от размножения старых, изношенных особей, которые могли бы воспроизвести лишь слабое потомство. Этим объясняется, что чем выше плодовитость животного, и чем менее истребляется его молодь, тем короче является его жизнь. Долголетие птиц связано именно с их малой плодовитостью. Эта последняя объясняется опять-таки биологически. Птицы не могут одновременно откладывать большое число крупных яиц, так как их значительный вес был бы большим препятствием при летании. Куриные откладывают еще значительное число яиц, которые, однако, легко становятся добычей врагов ввиду сравнительной доступности их гнезд. Голуби и хищники откладывают единовременно только 2 яйца, а буревестники даже 1 яйцо и притом несутся 1 раз в году. Так как и яйца и молодь птиц все же гибнут в заметном количестве от врагов или неблагоприятных случайностей, то необходимость оставить после себя число потомков, покрывающее эту гибель, достижима только значительным продлением жизни.
У млекопитающих плодовитость в общем выше, а процент гибели их молоди сравнительно низок; соответственно и жизнь млекопитающих короче (особенно, например, у большинства грызунов). Наоборот, животные с малой плодовитостью живут значительно дольше (человек, киты, слоны рождают по 1 детенышу, при чем последние не чаще 1 раза в 2 года). {51}
Продолжительность жизни половозрелого животного явно зависит и от способов откладки яиц, от того, существует ли охрана кладки, или другие виды заботы о потомстве, чему можно привести много примеров из различных типов животных.
Таким образом биологически продолжительность жизни оказывается связанной прежде всего с воспроизводительной способностью животного и биологическими особенностями воспроизведения и выведения потомства.
В данном случае мы имеем зависимость собственно одного только периода зрелости от указанных факторов. Что же касается вообще длительности отдельных периодов жизни, то и здесь наблюдаются известные закономерности. Так, у млекопитающих более продолжительный эмбриональный период и период юности соответствуют обычно и более продолжительному периоду зрелости, а следовательно и вообще более длительной жизни. Например у лошади эмбриональный период длится 11 месяцев и период юности 3—4 года; у крупного рогатого скота, общие размеры тела которого приблизительно те же самые, эмбриональный период длится 9 месяцев и период юности 1/2—2 года; соответственно и общая жизнь последних почти в 2 раза короче, чем у лошади.
Что касается длительности периода роста, то она вообще находится в большой зависимости от питания животного. В особенности это ясно у насекомых. Личинки мух, питающиеся гниющим мясом, и личинки пчел, питающиеся медом и пыльцей, достигают стадии куколки в несколько дней. Гусеницы бабочек, питающиеся менее питательной пищей — листьями, окукливаются через 1—2 месяца. Еще продолжительнее личиночный период у жуков, питающихся растительным перегноем, и в особенности велик он у короедов.
Индивидуальная жизнь взрослого животного может быть заметно продлена благодаря существованию периодов покоя, длящихся иногда в течение нескольких лет.
Можно думать, что и те периодические состояния покоя, которые наблюдаются у некоторых животных в виде явления сна, способствуют общему удлинению жизни уже потому, что в это время трата живого вещества значительно понижена; по всей вероятности, такое состояние покоя, способствуя более полному удалению из организма вредных продуктов обмена, как бы освежает организм и обеспечивает ему более длительное существование. Такой сон — отдых — связан всеми переходами с явлениями покоя, наблюдающимися нередко у животных при наступлении неблагоприятных условий {52} существования, как голод, холод, высыхание водоемов у водных животных и т. д. Таковы, например, зимняя спячка некоторых млекопитающих, которая является в сущности сном, нередко прерываемым краткими периодами бодрствования. При этом обмен веществ крайне понижен, что выражается и в значительном падении температуры тела. Зимний покой составляет правило для амфибий, рептилий, всех наземных и многих водных беспозвоночных. В этом случае мы имеем обычно полную остановку всех жизненных проявлений — так называемое состояние „скрытой жизни”. Состояние скрытой жизни может быть вызвано и высыханием водоемов (коловратки, тихоходки), или просто недостатком влаги (улитки), а также и голодом. Таким образом и голод, вызывая понижение обмена до минимума, может привести к значительному продлению жизни. Известно, что, например, обыкновенный постельный клоп может в таком состоянии покоя под влиянием голода пребывать целые годы.
Мы видели, что с течением жизненного процесса связано состаривание организма, которое у сложных животных всегда приводит к смерти. Причина смерти и срок ее наступления заложены в самой организации животного. Мы уже рассмотрели те процессы старения, которые наблюдаются с течением времени в организме, и высказали свои соображения об общих причинах смерти как явлении, связанном с ограничением роста животного и прекращением клеточных делений, по крайней мере в наиболее дифференцированных органах. Определенность организации, связанная с точным нормированием запаса живого вещества, ведет к установлению и определенного нормального срока жизни.
Нам остается подробнее ознакомиться с различными воззрениями на непосредственные причины старения и смерти.
Вейсманн полагает, что смерть есть полезное приспособление, приобретенное животными во время их филогенеза путем естественного отбора. По Вейсманну, смерть определяется появлением трупа, и так как последнего у простейших нормально не наблюдается, то простейшие бессмертны. Смерть приобретена многоклеточными животными одновременно с дифференцировкой на сому и половые клетки. Клетки тела — сома — утратили свое бессмертие, и непосредственной причиной их смерти является изнашивание в результате функциональной их деятельности. {53}
Гётте энергично восстает против взглядов Вейсманна. По Гётте, смерть не есть результат изнашивания и не есть приобретение многоклеточных организмов.
Смерть характерна для всех организмов, в том числе и для простейших. Смерть в определении Гётте есть прекращение жизни индивидуума. И простейшие умирают, но только момент смерти у них совпадает с размножением. Размножение и есть причина смерти. При всяком делении простейшего его индивидуальность теряется — оно умирает, хотя и без образования трупа, так как все его живое вещество идет на построение новых индивидуумов. Смерть сложного организма не есть результат изнашивания и гибели его клеток. Жизнь сложного организма едина и не является суммой функций отдельных частей. Строение организма есть лишь условие его жизни, и жизнедеятельность клеток является источником сил для работы всего живого организма. Смерть есть остановка общей жизни индивидуума. Причиной этой смерти Гётте считает и у сложных животных размножение. Образование половых клеток нарушает строение всего организма, и это нарушение рано или поздно и ведет к смерти.
Однако понятие о смерти становится совершенно расплывчатым и неопределенным, если вслед за Гётте считать за смерть всякое прекращение индивидуальной жизни. Не только деление, но и инцистирование простейших принимается за смерть. Является вопрос о других формах размножения, как, например, почкование. Является вопрос о периодах покоя и скрытой жизни, о таких даже явлениях, как патологическое раздвоение личности. Что же и это все смерть? Несомненно, что такое чрезмерное расширение понятия смерти не вносит ясности в спорную проблему, и нельзя закрывать глаза на то, что прекращение существования индивидуума при делении у простейших и смерть сложных животных представляют собой явления глубоко различные.
По Мечникову, в организме происходит непрерывная борьба частей. В результате различных несовершенств организации, как, например, существования обильной бактериальной флоры в кишечнике, происходит постепенное отравление организма (токсинами кишечника и другими ядами). При этом хроническом отравлении страдают прежде всего менее стойкие специализированные клетки — благородные ткани (мускульная, нервная). Это дает другим, менее специализированным клеткам — неблагородной, соединительной ткани,— перевес в борьбе. Соединительная ткань разрастается за счет {54} благородных тканей, гибнущих при активном участии фагоцитов. Фагоциты пожирают ослабленные нервные клетки и освобождают место для клеток соединительной ткани.
Позднейшие исследования подтвердили большое значение фагоцитоза при развитии старческих изменений в различных органах и в особенности в нервной системе, но выяснили несколько иную роль фагоцитов в этих явлениях. Фагоциты уничтожают не живые еще клетки, а исключительно уже погибшие — они являются в роли санитаров, очищающих организм от клеточных трупов и их продуктов распада. Таким образом их роль является, в сущности, обратной той, которую предполагал Мечников, — они способствуют не укорочению жизни, а ее продлению. Более того, в настоящее время имеются уже наблюдения, указывающие на еще более положительную роль фагоцитов. По Гармсу, фагоциты собираются у поверхности нервных клеток, перегруженных зернами старческого пигмента, и освобождают их от этих продуктов распада. Нервная клетка оказывается тогда освеженной, омоложенной.
Милльман видит причину смерти в ухудшении условий ассимиляции, связанном с тем, что при процессе роста организмов их масса растет пропорционально кубу диаметра, а поверхность — пропорционально квадрату. Таким образом в наихудших условиях оказываются органы, растущие всей своей массой, как, например, центральная нервная система. В относительно хороших условиях оказываются органы, увеличивающие главным образом свою поверхность, как кожа, кишечник. Наблюдения показывают, что такие органы действительно обладают наиболее продолжительным ростом.
Рубнер считает основной причиной старения и смерти прекращение роста. Только рост может устранить последствия одностороннего проявления жизни. При остановке роста и устранении возможности перестройки внутренней организации живое вещество постепенно теряет свою способность к ассимиляции.
Как уже указывалось, Вейсманн полагает, что смерть приобретена животными одновременно с дифференцировкой клеток животного на соматические и половые. Сома утратила свое бессмертие, так как жизнь целого организма нужна лишь до тех пор, пока не созрели и не отделились половые элементы. Непосредственной причиной смерти высшего животного является изнашивание некоторых тканей, быть может и неизбежное при их высокой дифференцировке, но эта последняя только потому и оказалась возможной, что бессмертие не имело никакого значения для жизни вида. {55}
Таким образом дифференцировка тканей и клеток организма не совместима с его бессмертием, но, в то время как по Вейсманну она является последствием утраты бессмертия, по Майноту она оказывается ее причиной. На основании анализа явления роста организма во время его индивидуального развития Майнот видит причину старения и смерти в нарастании и дифференцировке протоплазмы (цитоморфоз), в результате которой замедляются и, наконец, прекращаются клеточные деления.
По Чайльду, старение связано с понижением приспособляемости и понижением регенерационной способности, что доказывается главным образом опытами над планариями, регенерационная способность которых и приспособляемость к растворам алкоголя с возрастом непрерывно падает. По Чайльду, клеточный обмен приводит к образованию определенных структур; так как эти „структуры” состоят из химически более инертных веществ, то они по мере возрастания их массы являются все большим препятствием для самого обмена, который, наконец, оказывается недостаточным. Прогрессивное падение обмена и представляет собой самый характерный процесс, сопровождающий старение организма.
Однако продукты дифференцировки, или клеточные „структуры”, в зрелом возрасте уже заметно не возрастают в массе, и таким образом падение обмена взрослого организма вряд ли объяснимо с точки зрения Майнота и Чайльда. От чего же, однако, зависит это падение? Можно предположить, что организм появляется на свет с известным запасом внутренней энергии, которая постепенно расходуется (по Бюлеру, с известным запасом химического сродства к веществам, служащим питательными; ассимиляция, рост и развитие покоятся таким образом на постепенном сглаживании электрохимических различий и поэтому неизбежно приходят к концу). Быть может, как это теперь многими принимается, половые клетки нагружены известными энзимами, определяющими в дальнейшем характер обмена. Если эти энзимы растрачиваются и не возобновляются (Бючли), то это объясняло бы нам и падение обмена и связанное с этим неполное удаление его продуктов и другие явления старости. Жизненный процесс есть сложный химический процесс определенной длительности (Лёб). Однако несомненно, что организм может возобновлять такие запасы. Созревшие половые продукты оказываются нагруженными нормальным запасом потенциальной энергии в форме, быть может, энзимов или других веществ, получаемых в результате обмена в материнском организме. Таким образом можно лишь {56} предположить, что эта способность сохраняется не во всем организме, как целом, а лишь в известных его частях и притом, вероятно, главным образом в половых клетках; она обычно безвозвратно теряется в дифференцированных тканях тела. Последняя попытка разобраться в вопросах, связанных с проблемой старости и смерти принадлежит, насколько мне известно, Домсу (21)*). Домс справедливо отмечает неправильность попыток объяснить явления старости теми или иными возрастными изменениями организации. Всякие нарушения нормальной функции, всякие явления дезорганизации представляют собой не причины, а лишь симптомы старости. Причина старения лежит в тех моментах, которые препятствуют восстановлению разрушенного, и должна заключаться в общей конституции всех Сложных животных. Домс ищет эту причину в тонкой организации клетки и именно в ядре как носителе всех наследственных свойств организма. Домс развивает гипотезу, опирающуюся на теорию биобластов О. Гертвига. Так как при развитии зародыша несомненно ядра всех его клеток получают всю наследственную субстанцию, весь комплекс „ биобластов”, то процесс дифференцировки объясним только тем, что в различных клетках активируются, переходя, очевидно, частью в плазму, вполне определенные биобласты. Эти активные, функционирующие биобласты растут и размножаются весьма энергично. Полный набор биобластов, оставшихся в ядре в инактивном состоянии, также растет и размножается, но лишь постольку, поскольку происходят еще клеточные деления, сопровождающиеся их удвоением и распределением поровну между дочерними клетками. После законченной специализации и прекращения делений весь набор биобластов в ядре остается бездеятельным. Биобласты представляют собой индивидуумы со всеми элементарными жизненными проявлениями. Как и в любой части организма, функциональные раздражения и здесь ускоряют ассимиляцию, а отсутствие этого раздражения ведет к атрофии.
Хроническое отсутствие функционального раздражения, необходимого для нормального обмена, ведет ко все возрастающему повреждению инактивных биобластов. Так как жизнь целого обусловлена нормальным и гармоническим функционированием всех его частей, то хроническое повреждение отдельных биобластов ведет к хроническому нарушению обмена веществ в ядре и к ослаблению всей клетки. Таким образом старение есть следствие расхождения между полнотой набора наследственных зачатков и частичной их реализацией в отдельных частях организма сложных животных. У простейших старения нет, {57} так как все зачатки реализуются в нерасчлененном их теле. Действительной причиной старения, по Домсу, является, следовательно, не изнашивание, связанное] с функционированием, с активностью, а атрофия, связанная с отсутствием функции, с инактивностью. Возможность омоложения покоится на активировании инактивных биобластов при всякой регуляции. Половое размножение связано с особенно полным омоложением, так как в половых клетках активируется весь набор биобластов.
Интересная попытка Домса подойти к явлению старости с совершенно новой точки зрения опирается, как сказано, на теорию биобластов О. Гертвига. Эта теория, как и другие родственные теории, представляет собой, однако, далеко не доказанную гипотезу. И наоборот, в настоящее время накапливается все больше данных, указывающих, что наследственные единицы не являются представителем того или иного признака, ткани или органа, а влияют на всю сумму организации. Таким образом нужно думать, что инактивных биобластов вовсе и не существует — они все активны, хотя их деятельность может получить различное направление в зависимости от тех условий, в которые они попадают в данной клетке, ткани, органе. Не доказано кроме того, что биобласты можно рассматривать как жизненные единицы, столь самостоятельные в своей ассимиляционной деятельности, как этого требует гипотеза Домса. Однако помимо этой гипотетичности построений являются и другие затруднения. Гипотеза Домса не объясняет относительной стойкости клеток высших организмов, жизнь которых часто оказывается весьма длительной (птицы), между тем как увеличение общего числа инактивных биобластов в любой клетке сложно дифференцированного организма должно было бы привести у них к сокращению жизненного срока. Она не объясняет и удивительной стойкости наиболее специализированных клеток, в которых, очевидно, запас инактивных биобластов особенно велик (нервные клетки) и которые тем не менее бессменно функционируют в течение всей жизни.
Так как наибольшая степень омоложения достигается в половых клетках и, следовательно, не в результате функционального раздражения биобластов, т.-е. действия адэкватного раздражителя (как в специализированных клетках при их функционировании, когда нарастают активные биобласты в плазме), а в результате действия какого-то инадэкватного раздражителя, действующего на все биобласты (вызывая их рост и деление клеток), то, мне кажется, логичнее было бы рассматривать и старение как результат выпадения инадэкватного {58} раздражителя, вызывающего рост всей идиоплазмы, т.-е. клеточные деления (или, как мы здесь принимаем, наоборот, как результат воздействия фактора, тормозящего рост идиоплазмы, а следовательно и клеточные деления, при общем ограничении роста индивидуума).
Если рассматривать весь организм как целое, то для него, как указывалось уже раньше, характерна точная определенность жизненного цикла. Развитие организма начинается от яйца и не прекращается до самой смерти — весь жизненный цикл представляет собой длинную цепь характерных возрастных изменений. Смерть целого есть необходимое завершение жизни сложного животного — смерть есть последняя фаза жизни (Ферворн и др.).
Хотя смерть всего организма и не есть смерть отдельных клеток — многие клетки сложного организма являются потенциально бессмертными (Дофлейн, Дюркен), но все же смерть есть следствие изменений, происходящих в отдельных клетках (Шаксель, Жорес). Эти возрастные изменения имеют характер неизбежного строго предопределенного процесса. Первый период жизни (ранние стадии эмбрионального развития) характеризуется относительным нарастанием ядра, что может быть названо помолодением; второй, значительно более длинный, период — созревания и состаривания — характеризуется увеличением и дифференцировкой протоплазмы (цитоморфоз, Майнот). Дифференцировка соматических клеток возможна лишь при утрате способности размножения путем деления, т.-е. при утрате своего потенциального бессмертия (Вейсманн, Майнот). Односторонняя специализация клеток связана с неспособностью устранения дефектов, возникающих в результате их функционирования.
Вряд ли, однако, только дифференцировка сама по себе ведет к ограничению срока жизни. Смерть появляется впервые там, где нет еще дифференцировки между элементами сомы (колония Volvox) и нельзя сказать, чтобы жизнь выше организованных животных была короче жизни менее дифференцированных. Наконец, наиболее дифференцированные клетки организма фактически обладают ничуть не меньшим нормальным жизненным сроком, а, наоборот, даже большим, чем мало дифференцированные. Нервная или мускульная клетка живут в течение всей жизни, между тем как клетка эпидермиса позвоночных или кровяная клетка имеют самое кратковременное существование (первые не замещаются, между тем как вторые имеют заместителей). Наиболее специализированные клетки оказываются и наиболее стойкими при неблагоприятных условиях существования (например нервные клетки при голодании). {59}
Поэтому мы считаем причиной смерти не дифференцировку, а ограничение роста. Последнее достигается, очевидно, существованием некоторого внутреннего регуляторного аппарата в виде известного запаса энергии, растрачиваемого во время течения процессов роста и деления, а также в виде системы известных специфических веществ с характером тормозов, вырабатываемых самим организмом (гормонов)*). Что гормоны в действительности могут определять форму и даже поддерживать ее, не подлежит никакому сомнению; развитие вторичных половых признаков под влиянием гормонов половых желез является наиболее ярким примером такого формирующего воздействия.
Но каким образом ограничение роста, т.-е. отсутствие клеточных делений, ведет к смерти? Мы видели, что клетка без обновления своей организации, и в частности ядерного аппарата, не может функционировать вечно — она изнашивается. Изнашиваемые части в молодых клетках сначала успешно восстанавливаются, но затем с падением ассимиляции, связанным с ограничением роста, и регенерационная способность падает. Дефекты уже не устраняются полностью (известен факт упрощенной и недостаточной регенерации органов и тканей, то же, повидимому, касается и клеточных структур). Это изнашивание ведет прежде всего к несовершенному удалению продуктов обмена, благодаря чему последние накапливаются в теле клетки и вообще в организме (Гензен).
Продукты распада не представляют собой индиферентных тел, они являются ядами, и таким образом происходит постепенное самоотравление организма (Монтгомери). Во многих случаях такое накопление продуктов распада ясно обнаруживается в виде отложений старческого пигмента (Рёссле).
Таким образом смерть есть, в сущности, результат несовершенного обмена веществ в изношенных клетках. При более совершенной ассимиляции и постоянном восстановлении разрушаемых частей жизнь могла бы быть продлена, и высшего предела ее длительности установить невозможно.
Во многих органах клетки по мере изнашивания замещаются другими, но в наиболее специализированных частях организма, и в частности в центральном регуляторном аппарате, в нервной системе, эта смена невозможна без нарушений жизненных функций всего организма. В этих высокоспециализированных органах специфические клетки постепенно гибнут и замещаются разрастающейся соединительной тканью (склероз). {60}
Таким образом стареет прежде всего нервная система, а затем в связи с этим наступают нарушение питания и атрофия различных других органов, и в частности некоторых органов внутренней секреции, на что в последнее время обращено Особо значительное внимание.
Мы рассматриваем здесь атрофию некоторых органов внутренней секреции как явление вторичное. Некоторые авторы (Лоран) видят в них, наоборот, первичный процесс, который уже влечет за собой и иные явления старости. Это мнение основано на существовании у высших позвоночных характерных возрастных изменений инкреторного аппарата.
Для периода роста высших позвоночных характерны значительное развитие и деятельность зобной железы, которая стимулирует рост различных органов, в особенности рост костей и эпифизы (шишковидной железы), которая, очевидно, тормозит развитие половых желез и вторичных половых признаков. Эти железы испытывают затем обратное развитие, рост ослабевает, окончательно формируются половые железы, и в связи с инкреторной деятельностью последних развиваются вторичные половые признаки. Организм переходит в состояние зрелости. Последняя характеризуется преобладанием деятельности щитовидной и половой желез. К старости деятельность этих желез ослабевает, и они частью атрофируются. В этом возрасте весьма ясно выражена деятельность надпочечных желез. По Лорану, ослабление деятельности щитовидной железы влечет за собой ряд явлений старости (склероз, поседение и др.), а ненормальное усиление деятельности надпочечников вызывает артериосклероз.
Не подлежит никакому сомнению, что деятельность инкреторных желез имеет колоссальное значение в смене фаз жизненного цикла любого организма. Эти железы составляют часть того регуляторного аппарата, который нормирует формообразование и рост животного. Другие такие же железы регулирует обмен и другие жизненные функции организма и отдельных его частей. Такой бессменно работающий аппарат с высоко-специализированным, специфическим для каждого органа, обменом, подлежит, конечно, изнашиванию не менее, чем нервная система.
Однако вряд ли имеются данные приписывать этим органам руководящую роль в наступлении старческих изменений. Возрастные изменения наблюдаются вообще во всей организации, и мы знаем немало провизорных органов с весьма иногда кратковременным существованием. Видеть во всякой атрофии органа старческую {61} дегенерацию нет никаких оснований. Тем менее имеются основания искать источника явлений старости и смерти специально в атрофии какого-либо определенного органа внутренней секреции. Эта атрофия оказывает, конечно, немалое влияние на течение жизненных процессов, способствуя их полному расстройству точно так же, как и явления старческой дегенерации в нервной системе, сосудах и других жизненно необходимых органах. Но все же, повидимому, атрофия половых желез, щитовидной и пр. есть не причина, а просто одно из выражений старческих изменений организма, если даже не их результат. Усиление секреции надпочечников представляет, быть может, компенсацию старческого ослабления симпатической нервной системы. Если же дегенерация последней, быть может, связана с надпочечниками, то лишь в той же мере, в какой связана и их нормальная функция. В этом случае надпочечники представляют лишь часть общего регуляторного аппарата, рассчитанного на определенную продолжительность жизни. Отравляющее действие надпочечников, как и все самоотравление организма, есть лишь выражение несовершенного обмена, который с течением времени сказывается все сильнее.
Повторяя наши выводы, мы можем их резюмировать следующим образом.
Ограничение жизненного срока введением смерти есть результат ограничения роста и формы животного определенными нормами. Это ограничение достигается существованием внутреннего аппарата, устанавливающего предел клеточным делениям и рассчитанного иногда даже на определенное число клеточных делений. Этот внутренний аппарат может быть внутриклеточным — например в виде известного запаса энзимов, расходуемых при последовательных делениях, и может быть внеклеточным в виде специфических гормонов, заторможивающих клеточные деления*).
Прекращение клеточных делений совпадает с клеточной дифференцировкой, и наиболее дифференцированные клетки окончательно теряют способность делиться. Однако дифференцировка не есть единственная причина прекращения клеточных делений, так как нередко последнее наблюдается и в совсем мало дифференцированных тканях. Клеточная дифференцировка не ведет прямо к укорочению жизненного срока. Наоборот, совершенное разделение труда и специализация, связанные с более совершенным выполнением жизненных функций, в том числе и с более совершенным возобновлением разрушенных частей, ведут к увеличению стойкости и продолжительности {62} жизни. Все же смерть неизбежна, так как жизнедеятельность неуклонно падает, и наступают явления старческой дегенерации. Эти изменения мы можем рассматривать как неизбежное следствие ограничения роста. Ограничение роста есть ограничение ассимиляции. Клетки, утратившие способность роста и деления, в значительной мере утратили и способность синтеза своего специфического живого вещества. Их протоплазма сгущается. Они слабо регенерируют и не в состоянии полностью устранять последствия случайных или хронических (функциональных) повреждений. Постепенно накапливающиеся дефекты внутренней организации клеток все более отзываются на их функции и на деятельности всего организма. Первым признаком нарушения жизнедеятельности клеток является недостаточное удаление продуктов распада. Таким образом старческие изменения представляют собой результат нарушения обмена в изношенных клетках, благодаря чему происходит накопление продуктов распада.
Полное обновление внутренней организации клетки возможно только при полном восстановлении ассимиляционной деятельности, как она выражается прежде всего в нарастании живого вещества и в наличии клеточных делений. Однако и при прекращении последних можно себе представить почти беспредельное увеличение нормальной продолжительности жизни при условии идеально совершенной клеточной ассимиляции, при которой происходит полное восстановление живого вещества, поддерживающее внутреннюю гармонию клеточной организации, и при точной регуляции обмена во всем организме.
Если старение и смерть есть в конце концов результат совершенно необходимого ограничения роста организма, достигаемого существованием лишь определенного запаса потенциальной энергии и существованием особого внутреннего аппарата, регулирующего явления роста и клеточных делений, то не возможно ли омоложение путем устранения тормозящих гормонов, путем возобновления запаса энергии, или, по крайней мере, путем нового возбуждения ассимиляционной деятельности действием каких-либо специфических гормонов?
Все это оказывается возможным, хотя и не всегда в пределах индивидуума.
1. Устранение тормозящих гормонов при изолировании небольших частей организма и культивировании их вне последнего (эксплантация) приводит к неограниченному размножению, по крайней мере, некоторых „бессмертных” клеток. Тоже самое наблюдается и в пределах организма под влиянием невыясненных еще причин, когда {63} физиологическая изоляция известных частей приводит к образованию опухолей с неограниченным ростом.
2. Дедифференцировка известных частей организма под влиянием повреждений, голода или других не изученных еще влияний может в известных условиях привести к возобновлению запаса внутренней энергии и к восстановлению утраченной части или всего организма (реституция и воспроизведение вообще в различных его видах).
3. Наконец, специфические гормоны могут обновить весь организм и также поднять и его регенеративные способности.
Мы уже раньше указывали, что смерть целого организма не есть результат умирания его частей, его клеток.
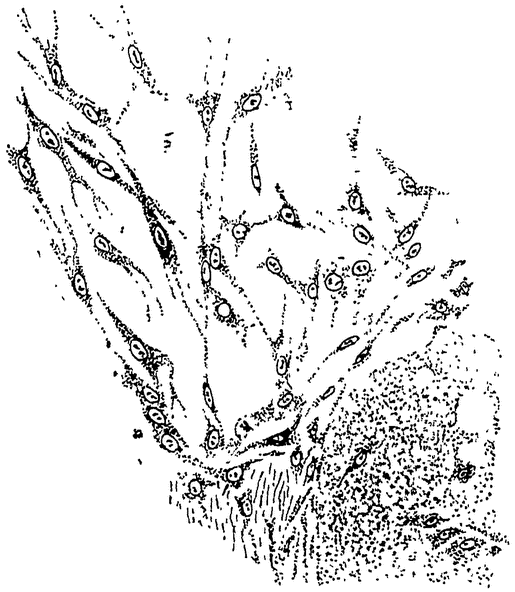 |
Рис. 20. 39-дневная культура соединительной ткани кролика. Видны размножающиеся клетки (фибробласты). По Максимову. |
Еще интереснее так называемые тканевые культуры. Незначительные кусочки тканей, стерильно помещенные в питательную среду (кровяную плазму или искусственные среды) при естественной температуре тела не только остаются живыми в течение продолжительного времени, но и растут при деятельном размножении их клеток. Особенно быстро растут различнейшие эмбриональные ткани. Медленнее размножаются клетки в тканевых культурах, полученных от взрослых животных. Наиболее дифференцированные клетки последних, как нервные и мускульные, вовсе не делятся. Энергичнее всего размножаются клетки в культурах соединительной ткани (рис. 20). Эти последние при постоянной смене питательной, среды („пересеве” во избежание {64} накопления продуктов обмена) могут поддерживаться месяцы и годы. Повидимому, такие культуры бессмертны, так как до сих пор не удалось подметить даже падения скорости клеточных делений.
Ясно, однако, видна разница в скорости деления клеток в культурах от молодых и от старых животных. В последних клетки делятся гораздо медленнее. Таким образом клетки животного не молодеют в тканевых культурах, но и не стареют. Старость есть явление, развивающееся именно в организме (в результате ли падения ассимиляционной деятельности специализированных клеток органов ограниченного роста или под влиянием постоянного воздействия гормонов, тормозящих клеточные деления). Вне организма дальнейшего старения не наблюдается. Это следует поставить в связь с существованием клеточных делений как фактора, освежающего отдельные клетки и, по крайней мере, как показателя интенсивности ассимиляции, которая, между прочим, покрывает и всю трату вещества.
Здесь ясно видно, что бессмертие клеток связано именно с сохранением способности размножения. Клетки, утратившие под тем или иным влиянием свою способность к росту и делению, всегда стареют и умирают.
Подобный же неограниченный рост тканей при непрерывном размножении клеток наблюдается и в организме патологически при образовании злокачественных опухолей. В пользу того, что и здесь имеет место какая-то „физиологическая” изоляция известного участка тканей, говорят новейшие эксперименты Шакселя, который перевивал обратно на организм тканевую культуру, полученную из регенерационной ткани у аксолотля и получал при этом образование опухолей неограниченного роста.
Таким образом ясно, что не все соматические клетки смертны сами по себе, но в организме они находятся под влиянием воздействий, подавляющих их обмен, их клеточные деления и тем самым вызывающих их старение.
Только специализированные клетки теряют свою способность к делению по внутренним причинам и вместе с тем утрачивают свое бессмертие.
При известных условиях животные ткани могут подвергнуться процессу обратного развития или дедифференцировки. Это наблюдается, например, нередко при воспалительных процессах, при {65} поранениях, при пересадках и в тканевых культурах. Такое обратное преобразование может привести данную ткань или клетки к гибели, но может закончиться и их восстановлением, новой дифференцировкой и омоложением известной части или даже всего организма.
Омоложение обусловлено в этом случае возобновлением клеточных делений. Наблюдаемое размножение клеток связано, повидимому (Габерландт), с появлением известных раздражителей — гормонов, действующих как возбудители клеточных делений. Такие гормоны появляются, по всей вероятности, в особенности в результате поранений. Их действие обнаруживается, однако, не только в поврежденных, но и в соседних тканях, где также наступают клеточные деления.
Таким образом живой организм реагирует на раздражение при всяком поранении восстановлением поврежденной части тела — регенерацией. Регенерационная способность представляет собой основное свойство всех живых организмов, которая, однако, проявляется у разных животных в различной степени. Иногда дело ограничивается лишь образованием рубцевой ткани, затягивающей рану (обычно у высших позвоночных), в других случаях таким образом могут восстанавливаться целые утраченные органы (например конечности или хвост у рыб и у земноводных), но в некоторых случаях путем регенерации может быть восстановлена значительная часть всего организма, иногда из ничтожнейшей его части.
Высокая регенерационная способность обычно связана с низкой организацией, т.-е. со слабой дифференцировкой тканей. Так, например, чрезвычайно высока эта способность у кишечнополостных и притом особенно у гидроидных полипов. Обыкновенная гидра представляет собой классический объект, на котором издавна производились разнообразнейшие эксперименты подобного рода. Не менее высока регенерационная способность и у ресничных червей (рис. 21). Различные виды планарии также способны восстановить все свое тело из ничтожнейшего куска. Однако и такие высоко-организованные животные, как кольчатые черви, немертины и асцидии способны заново перестраивать всю свою организацию и восстанавливать ее из небольших частей. У кольчатых червей, с относительно очень сложно построенным организмом, очень незначительная часть тела, иногда даже 1 сегмент, может вновь воспроизвести целого червя в составе 100—150 сегментов (Lumbriculus). В этом случае, впрочем, в организме имеются запасы эмбриональной ткани, за счет которой и идет регенерация. Последняя у многих червей является {66} способом размножения. В тех случаях, когда последнего нет, резервных тканей обычно не имеется, и регенерация идет из остатков старой ткани.
Если характер повреждения таков, что нарушается нормальное питание организма, что бывает обычно при удалении части тела у червя, то регенерация утраченной части идет непосредственно за счет старых тканей организма, большая часть которых при этом гибнет.
При регенерации отдельные ткани развиваются обычно за счет сохранившихся остатков ткани того же рода. Если же орган или
 |
Рис. 21. Регенерация у планарий. В и С — регенерация из двух половин тела (A); D — регенерация из небольшого отрезка туловища; Е — регенерация из головной части; F и G — регенерация целых планарий из небольших отрезков головы. По Моргану. |
часть тела удалены целиком, то наблюдается всегда „метаплазия”, т.-е. сначала дедифференцировка некоторых клеток и тканей, преобразовывающихся в индиферентную ткань эмбрионального характера, а затем новая дифференцировка уже в ином направлении.
Во многих случаях регенерации органов трудно бывает установить с несомненностью процесс дедифференцировки вполне дифференцированных, клеток. Во многих органах нередко сохраняются запасы индиферентных клеток, служащих для нормальной замены функционирующих и изнашивающихся тканей. При регенерации появляется всегда более или менее значительная масса эмбриональной ткани, источник которой не всегда можно установить, и которая во многих случаях могла бы получиться путем последовательного {67} деления отдельных резервных клеток. В некоторых случаях, однако наличие дедифференцировки доказано с несомненностью.
Классическим примером такой де- и передифференцировки является регенерация хрусталика хвостатых амфибий. Изученная первоначально Вольфом у тритона, регенерация хрусталика глаза протекает совершенно отлично от эмбрионального развития. В то время как эмбрионально хрусталик развивается в качестве утолщения эпидермиса, при регенерации он восстанавливается из совершенно иного источника — за счет края радужины (рис. 22). Радужина глаза состоит из вполне дифференцированных пигментных клеток. Каких-либо резервных клеток здесь не обнаружено. Клетки эктодермального эпителия радужины теряют свой пигмент, их ядра относительно увеличиваются
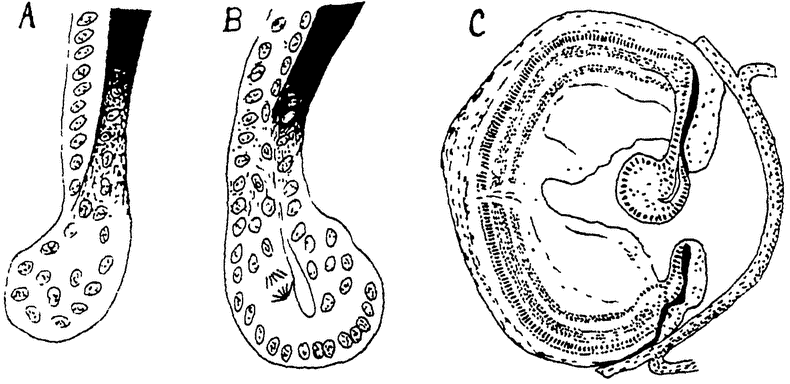 |
Рис. 22. Регенерация хрусталика глаза у тритона. A и В — край радужины с регенерирующим хрусталиком; С — весь глаз с регенерировавшим хрусталиком. По Вольфу из Моргана. |
и таким образом клетки принимают эмбриональный характер. Из этих омоложенных клеток формируется новый хрусталик, при чем составляющие его элементы испытывают новую совершенно определенную дифференцировку, превращаясь в прозрачные волокновидные клетки, расположенные строго определенным образом.
У кольчатых червей, как и у других животных, нервная система эмбрионально развивается из эктодермы. При регенерации органы и ткани обычно восстанавливаются по возможности за счет остатков соответствующих органов и тканей. Нервная система, однако, оказывается слишком специализированной и обычно обладает лишь ничтожной регенерационной способностью. Поэтому у кольчатых червей (у многощетинковых, по Е. Шульцу) при регенерации брюшной нервной цепочки эктодермальные клетки покровов дедифференцируются, выползают внутрь тела и превращаются в ганглиозные. У плоских червей нервная система при регенерации образуется {68} заново из паренхимы. Точно так же и у мшанок при регенерации нового полипида старые, очень плоские и бедные протоплазмой, эктодермальные клетки, вновь приобретают эмбриональные свойства.
Иногда, повидимому, и половые клетки могут образоваться путем дедифференцировки из соматических (у кишечнополостных, у турбеллярий и у олигохет).
Наконец, по мнению некоторых авторов (Рибберт, Ганземанн и др.), и злокачественные опухоли с неограниченным ростом (саркома, карцинома и др.) представляют собой результат дедифференцировки соматических клеток под влиянием каких-то раздражителей. Такие клетки могут с полным правом быть названы „бессмертными”, так как их культуры при перевивках на другие особи могут жить, очевидно, неограниченно долго1).
Во всех этих случаях дедифференцировка сопровождается восстановлением способности клеток к интенсивной ассимиляции и к размножению путем деления, т.-е. по существу может быть обозначена как омоложение.
Дедифференцировка не всегда, конечно, идет так далеко, как в приведенных примерах. Прежде всего возможность передифференцировки, повидимому, всегда остается в пределах зародышевых листков, т.-е. эктодермальные, например, органы регенерируют за счет эктодермы2). В большинстве случаев, однако, ограничение это идет гораздо дальше, и отдельные ткани и органы регенерируют только от уцелевших остатков соответствующих тканей и органов. Так, при регенерации конечностей у амфибий прежде всего рана затягивается надвигающимися с ее краев клетками эпидермиса, образующими наружные покровы регенерата. Затем под этим покровом образуется скопление эмбриональных клеток, в котором затем дифференцируются зачатки скелета, в непосредственной связи с остатками старого скелета, зачатки мышц, кровеносные сосуды, и врастают нервы путем регенерации от обрезанных концов старых нервов. {69} Эмбриональные клетки, составляющие основу регенерата, образуются путем дедифференцировки от равнозначащих тканей. Скелетогенная бластема образуется путем выселения и размножения клеток перихондрия или периоста, т.-е. от наиболее индиферентной (резервной) ткани прежнего скелета. Миогенные клетки образуются от обрезанных старых мышц путем дедифференцировки. В каждом мускульном волокне, как известно, можно различить существенную, специализированную его часть — совокупность сократимых мускульных фибрилль — и остатки недифференцированной протоплазмы с ядрами и митохондриями.
 |
Рис. 23. Перерождение и регенерация мускульного волокна после поранения. Последовательные стадии. По Циглеру из Майнота. |
Многие авторы и в этом и в других случаях отрицают существование явления дедифференцировки, так как здесь нет обратного развития дифференцированных частей протоплазмы, а имеются их гибель и восстановление за счет индифферентных частей протоплазмы. Это по существу верно; настоящего обратного развития здесь нет. Однако все же можно говорить о дедифференцировке, так как в результате этих процессов мы получаем ткань эмбрионального характера на месте и за счет прежней дифференцированной ткани (хотя бы и ценою гибели наиболее дифференцированных ее частей).
При значительных повреждениях явления дедифференцировки могут захватить и весь организм, в особенности в тех случаях, {70} когда нарушается его питание, и восстановление нового целого возможно лишь за счет старых тканей, потребляемых частью даже просто в качестве питательного материала. Такая „редукция” всего организма, ведущая затем к его „реституции”, наблюдается и не только при значительных механических повреждениях, но иногда и при других неблагоприятных условиях существования, как, например, особенно при интенсивном голодании.
У губок при ненормальных условиях существования наблюдалась редукция вплоть до образования отдельных круглых „редукционных тел” из однородных клеток (при лишении известковых губок солей
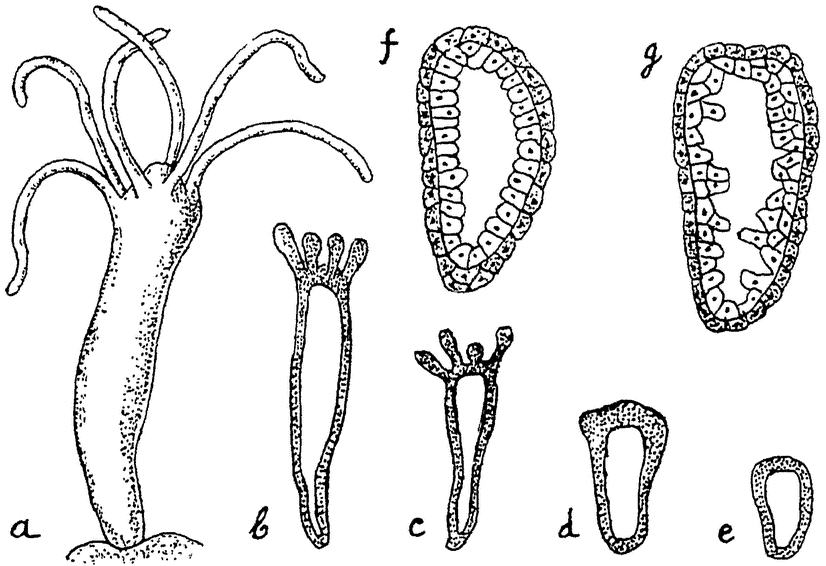 |
Рис. 24. Последовательные стадии редукции гидры при голодании (b — е); g — разрез через редуцированную гидру; f — разрез через нормальную стадию развития. По Бернингеру из Коршельта. |
извести, по Маасу), напоминающих почечки („геммулы”) пресноводных бадяг, которые образуются у них нормально к началу зимы. По Вильсону и Мюллеру, тело пресноводных губок можно растереть и просеять через густую ткань для изоляции отдельных клеток. Часть последних гибнет, другие сближаются путем амебоидных движений и образуют аггрегаты, из которых затем дифференцируются новые молодые губки. (По Гёксли, возможность передифференцировки ограничивается и здесь, однако, пределами зародышевых листков).
Гидроидные полипы также редуцируются при неблагоприятных условиях, после чего вновь восстанавливаются отдельные гидранты из уцелевшего материала. Экспериментально ход редукции прослежен особенно у пресноводной гидры в результате голодания {71} (рис. 24). В этом случае гидры постепенно уменьшаются в своих размерах, затем щупальцы укорачиваются и совершенно резорбируются. Получается гидра, лишенная щупалец (очень сходная с Protohydra). Рот зарастает, а тело гидры получает сначала булавовидную форму, а затем вид округлого двуслойного пузырька, вполне сходного с эмбриональной стадией планулы. Почкование прекращается уже в самом начале голодания, а половые клетки, наоборот, ускоренно созревают (Шульц, Бернингер). Значительное уменьшение размеров тела сопровождается гибелью большого числа клеток и прежде всего наиболее дифференцированных клеток, каковы, например, нематобласты. Отдельные клетки, повидимому, дедифференцируются и приобретают эмбриональный характер.
Голодающие планарии также весьма значительно сокращаются в размерах, так что объем их может уменьшиться до 1/300 первоначального. И здесь при таком интенсивном голодании наступает редукция. Значительная часть клеток и тканей и даже целые органы гибнут. Прежде всего разрушаются части сложного у планарии полового аппарата (но не сами половые клетки). Затем разрушаются и другие дифференцированные части за исключением нервной системы. Остаются наиболее эмбриональные ткани, выживающие за счет гибели специализированных частей организма (Шульц).
При значительных повреждениях, естественно ведущих к длительному голоданию (например продольный разрез тела), наблюдаются ярко выраженная дедифференцировка тканей и полная редукция организма и у немертин (К. Давыдов). Особенно ясно здесь дедифференцируется мускулатура, образующая тогда однородные группы совершенно эмбриональных клеток с крупными ядрами. Весь организм возвращается к эмбриональному состоянию. Наконец, классическим примером редукции сложного организма, ведущей затем к его восстановлению, являются некоторые асцидии (Clavellina и др.), как это впервые установлено Г. Дришем, а затем проверено многими другими авторами (Е. Шульц, Шаксель).
И вполне взрослые асцидии обладают очень высокой регенерационной способностью и при удалении даже значительных частей тела образуют регенерационную почку, за счет которой восстанавливается утраченная часть. Более молодые асцидии, в особенности при общих неблагоприятных внешних условиях, реагируют обновлением всего организма и при значительных повреждениях испытывают почти полную редукцию, т.-е. общее возвращение к эмбриональному состоянию (точнее к состоянию столониальной {72} почки). При наибольшей редукции асцидия превращается в небольшой комок, состоящий из наружного эпителия, с массой мезенхимы, в которой помещается зачаток кишечника и перибранхиальной полости. При редукции гибнет масса дифференцированных тканей и органов при деятельном участии фагоцитов (Шаксель). По новейшим исследованиям Шакселя, в этом процессе разрушения пощаженными остаются только комплексы индиферентных клеток. Как и у других животных, размножающихся вегетативным путем, в организме асцидий имеются значительные резервы индиферентных клеток. Шаксель приписывает последним очень большую роль и вообще при регенерации. В этом случае нет собственно дедифференцировки тканей, но дедифференцировка организма все же совершенно ясна. У некоторых асцидий в виде вполне регулярного явления наблюдалась также и ежегодная физиологическая редукция, с последующей реституцией организма (Кольери).
Чайльд наблюдал физиологическую редукцию и реституцию у стареющих планарий (Planaria velata). Задняя часть тела при этом расширяется и принимает форму диска; она теряет всякую активность и, наконец, отбрасывается; передняя часть тела восстановляет затем весь организм путем регенерации. Если задняя часть тела не отбрасывается, то организм зачастую гибнет. Но иногда происходит постепенная переработка материала, значительная часть совершенно рассасывается, но в конце концов весь организм перестраивается и возобновляется за счет дегенерировавших частей, потребляемых как питательный материал. Здесь, значит, в естественных условиях, после известного периода депрессии, после начавшегося разрушения, которое нередко распространяется на весь организм и приводит его к гибели, наблюдается в иных случаях полная реорганизация, полное омоложение. Является, однако, вопрос, представляет ли дедифференцировка организма или его части, действительно, омоложение, т.-е. ведет ли она действительно к продлению нормального срока жизни.
Соответственные эксперименты производились неоднократно. Чайльд (11) установил в результате тщательного исследования, что планарий, полученные путем регенерации из небольших частей тела, оказываются более жизнеспособными и устойчивыми, чем старые животные, от которых они произошли. Хотя при регенерации далеко не все тело развивается путем новообразования, процесс омоложения при этом охватывает все тело, т.-е. и старые ткани. Таким образом происходит полная реорганизация, полное обновление всего организма. {73} То же самое, по Чайльду, замечается и у кишечнополостных. Гармс (12) достигал продления жизни у кольчатого червя Hydroides, отделяя стареющую заднюю часть тела, которая затем возобновлялась путем регенерации. В новейшее время подобные эксперименты производились Гартманном над инфузориями (Stentor) и над ресничными червями (Stenostomum). У этих животных удавалось совершенно предотвратить размножение делением постоянной регенерацией отрезываемых частей тела. По Гартманну, здесь путем периодического искусственного уменьшения системы достигается периодическое омоложение. Такие же эксперименты производились и над гидрами. Периодическая регенерация отрезанной подошвы освежала весь организм, подавляя вместе с тем и почкование и половое воспроизведение (Гётш).
Таким образом, если старость есть следствие ограничения ассимиляции и прекращения клеточных делений, то, наоборот, возобновление последних при регенерации ведет к омоложению организма. В этом случае омоложение есть следствие известного насильственного воздействия, результат более или менее значительного повреждения организма. Регенеративные процессы наблюдаются в природе, однако, не только в результате насильственных повреждений организма. Иногда в ответ на различные более или менее значительные раздражения организм сам реагирует отбрасыванием известной части тела. Это явление, известное под названием автотомии, во многих случаях, у свободно передвигающихся форм, является приспособлением для освобождения от напавшего врага ценою гибели некоторой части тела (известный пример ящерицы, оставляющей свой хвост в руках ловца). У сидячих форм, однако, также иногда наблюдается автотомия, которая здесь вряд ли может иметь иное значение, чем физиологическое отбрасывание изношенных частей организма.
Автотомия производится животным путем сильного сокращения мышц чаще всего в определенных местах тела, которые оказываются специально для этого приспособленными. После отбрасывания органа или части тела наступает регенерация.
Отбрасывание отдельного органа наблюдается в особенности часто у членистоногих (у краббов, пауков и насекомых). В этом случае автотомируют конечности, отбрасываемые сокращением мышц, на определенном месте, отмеченном снаружи кольцеобразным утончением хитина в проксимальной ее части. При этом в соответствующем месте имеется иногда и специальная клеточная перегородка, {74}
 |
Рис. 25. Регенерация целой морской звезды из одного отделенного луча. Из Коршельта и Гейдера. |
Очень легко автотомируют морские звезды. Путем сильного сокращения кольцевой мускулатуры, они отбрасывают один или несколько лучей, после чего не только последние восстанавливаются, но и отброшенные лучи могут путем регенерации дать начало новым звездам (рис. 25). Автотомия наблюдается и у мшанок, легко отбрасывающих свои головки, а также и у моллюсков.
В особенности интересна автотомия у ресничных червей и у малощетинковых кольчатых. Здесь происходит отделение более или менее значительных частей тела, при чем каждая часть может восстановить затем организацию целого животного. В этом случае автотомия ведет к размножению животного путем особого рода быстрого и энергичного деления. Здесь омоложение связано, следовательно, с размножением.
Регулярное размножение сложных животных путем поперечною деления обыкновенно наблюдается в тех же группах червей (ресничных и кольчатых), в которых наблюдается и автотомия целых частей тела. Оба явления связаны между собой целым рядом промежуточных форм.
Если автотомия есть реакция организма на известное внешнее раздражение, то деление животного происходит главным образом вследствие известных внутренних причин, при чем внешнее раздражение играет лишь самую незначительную роль.
Таким, образом наблюдается, например, размножение делением на отдельные части у некоторых кольчатых червей (рис. 26). Каждая часть после разделения регенерирует целое животное. В некоторых случаях тело животного распадается на большое число мелких частей, и иногда даже обособляются отдельные сегменты (рис. 26), {75} которые регенерируют затем впереди новый головной конец, а позади — хвостовой.
 |
Рис. 26. Деление червя (Ctenodrilus monostylos). По Цеппелину из Коршельта и Гейдера. |
Обычно размножение делением отличается от автотомии тем, что в этом случае регенерация начинается раньше разделения, по крайней мере, появлением известной регенерационной зоны. У ресничных червей (некоторых планарий и у микростом) регенерация начинается задолго до деления, так что предварительно развиваются все недостающие органы обеих новых особей. Кроме того деления могут так быстро следовать друг за другом, что еще до конца первого разделения начинается новое деление обеих дочерних особей и т. д., так что в некоторых случаях таким образом развивается длинная цепочка молодых особей.
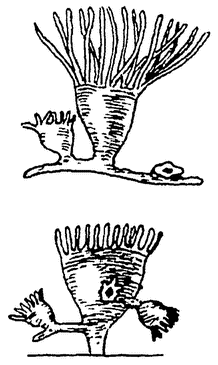 |
Рис. 27. Размножение сцифистомы путем образования столона и боковым почкованием. По Сарсу из Коршельта и Гейдера. |
Совершенно то же самое наблюдается и у кольчатых червей, у которых также наблюдаются нередко такие временные цепочки особей (Chaetogaster). У других кольчатых червей регенерационная зона развивается на заднем конце тела и дает начало длинному ряду молодых особей, являющихся как бы в виде придатка на теле материнской особи (Myrianida, Nais proboscidea).
К такому размножению тесно примыкают уже и другие виды бесполого размножения (рис. 27). У гидроидных полипов могут обособляться округлые или овальные тела, которые затем дают начало новым особям и колониям (фрустуляция). У актиний от подошвы развивается отросток, на котором затем появляется в виде выступа зачаток новой особи (фрагментация). У туникат на подобных же отростках (столонах) образуются почки, дающие начало новым особям, остающимся в связи со старой особью для образования колоний. {76} Наконец, сюда же примыкает и размножение путем почкования у гидроидов, кораллов и мшанок. У последних, кроме того, образуются еще особые внутренние почечки, состоящие из группы индиферентных клеток (эктодермальных и мезодермальных) — статобласты.
Во всех этих случаях почки образуются при самой незначительной затрате материала материнского организма. Только эта незначительная часть организма испытывает, следовательно, омоложение. Материнский организм сохраняет при этом свою индивидуальность, но зато он не избегает процесса старения и физиологической смерти.
Поэтому здесь нельзя говорить об омоложении особи, а лишь о возникновении новой молодой особи за счет незначительной части старого организма.
Отсюда уже один только шаг и до полового воспроизведения, которое представляет собой такой же процесс возникновения новой особи за счет минимальной части материнского организма, за счет одной только половой клетки.
И обособление половых клеток из общей массы клеток тела в некоторых случаях может быть отмечено процессом дедифференцировки. Именно для низших групп (губок и кишечнополостных) характерно позднее обособление половых элементов, и этот способ их образования, быть может, является первичным (Е. Шульц). В таком случае ранняя детерминация половых клеток, наблюдающаяся у большинства прочих животных, является процессом вторичным, вполне аналогичным образованию резерватов эмбриональных клеток у животных с высокой регенерационной способностью. Наконец, зрелые половые продукты и вообще являются специально дифференцированными клетками. В протоплазме яйца наблюдается нередко совершенно определенная дифференцировка, и, кроме того, она набита определенными запасными веществами. Сперматозоид является во всяком случае высоко специализированной клеткой. Эта специализация утрачивается во время оплодотворения, при чем дифференцированные части сперматозоида погибают. Таким образом и здесь имеется налицо ясная дедифференцировка клеток, предназначенных для восстановления целого организма. Если дифференцировка половых клеток возникает под влиянием материнского организма, то, быть может, дедифференцировка и здесь, как и во многих других случаях (эксплантация, трансплантация), первоначально есть результат нарушения связи с организмом. {77}
Было высказано мнение, что омоложение есть результат механического повреждения яйца сперматозоидом при процессе оплодотворения. Гормоны, возникающие при этом ранении, как и при регенерации, действуют, как возбудители клеточных делений (Габерландт). С этой точки зрения, однако, совсем необъяснимо явление партеногенеза.
Во всяком случае, следовательно, и половое размножение можно рассматривать, как результат дедифференцировки и омоложения известной, весьма небольшой, правда, части организма, за которой следует восстановление целого. Половое воспроизведение есть лишь, частный случай редукции и реституции (Чайльд).
Таким образом можно сказать, что в непрерывном процессе старения, неизбежно сопровождающем жизнь высших индивидуальностей, единство живого поддерживается периодическим омоложением, т.-е. дедифференцировкой более или менее значительной части индивидуума с последующим восстановлением всей организации в виде новой особи. Таким образом в жизни каждой особи кроется, по крайней мере незначительная, частица бессмертия.
Мы уже несколько раз коснулись вопроса о значении индивидуальности. Теперь нам остается рассмотреть этот вопрос ближе.
В случае размножения путем деления на две равные части, как: это наблюдается в особенности у простейших, индивидуальность. животного явно теряется одновременно с актом размножения.
Если деление происходит на неравные части, то здесь уже дается известный простор субъективным толкованиям, и только в случае резкого различия, как при типичном почковании, когда обособляется лишь незначительная часть организма, можно было бы уже определенно говорить о продлении существования индивидуума и о его независимости от размножения.
Новую, высшую и более устойчивую форму индивидуальности мы встречаем у колониальных простейших. Продолжительность жизни отдельных особей колонии оказывается при этом значительно увеличенной. В конце концов индивидуум гибнет, однако, вследствие изнашивания его сомы. Это изнашивание мы ставили в связь с невозможностью обновления ядерного аппарата при наблюдающемся здесь прекращении деления отдельных особей, колоний. {78}
Последнее явление, в свою очередь, связано с ограничением размеров колонии, что определяется уже комплексом биологических факторов. У сложных животных это выражено еще яснее.
Ясно, что развитие стойкой индивидуальности обусловлено установлением точно определенных соотношений с внешней средой. Биологические требования последней касаются, между прочим, и ограничения роста и определенности формы, как наружно видимого выражения такого приспособления. Организмы с неограниченным ростом совершенно немыслимы в реальных условиях существования. Такое ограничение естественно связано с прекращением клеточных делений, так как при постоянстве размеров клеток, характеризующих данный организм, рост обусловлен именно существованием клеточных делений. Последние связаны, как мы знаем, с перестройкой ядерного аппарата, которая, очевидно, играет роль омолаживающего фактора. Клетки, утратившие способность деления, всегда в конце концов изнашиваются и умирают.
Таким образом именно ограничение роста сложных животных ограничивает и жизнь известными пределами. Если бы произошло настоящее омоложение всего тела, т.-е. всех клеток такого организма, то это означало бы восстановление интенсивного обмена веществ, а вместе с тем и преобладание ассимиляции над диссимиляцией, столь характерное для молодого организма, т.-е. нарастание живого вещества, клеточные деления и неизбежный рост всего организма.
Мыслимо еще частичное омоложение известной части организма. Такое омоложение привело бы к образованию некоторой зоны роста, изменило бы общую форму и строение организма и тем нарушило бы его нормальные, жизненно важные соотношения с внешней средой. У животных со слабо выраженной и мало активной индивидуальностью, т.-е. не вполне фиксированными соотношениями с внешней средой, такое возобновление роста и изменение формы еще возможно.
Это касается, например, низших сложных животных — губок, форма тела которых во многих случаях оказывается непостоянной и рост не вполне ограниченным.
Гораздо чаще нарушение формы вследствие появления новых зон роста наблюдается в качестве временного явления. Вслед за нарушением формы следует ее восстановление путем регуляции — происходит обособление известной части организма в области зоны роста, и каждая часть восстанавливает форму целого организма. Если зона роста лежит посреди старых частей организма и разделяет {79} его на приблизительно равные части, то говорят о делении. Если зона роста выдается за пределы старого организма, и обособляется только это новообразование, то говорят о почковании.
В этих случаях омоложение и возобновление роста связано опять, как и у простейших, с увеличением числа особей, т.-е. с размножением организма. И здесь приходится повторить то же, что сказано о размножении простейших. Если при почковании материнский организм продолжает существовать и после обособления ряда новых особей, то при делении его индивидуальность теряется, растворяясь в произведенном потомстве.
Очевидно, и в этих случаях индивидуум есть образование не вполне стойкое. Может быть это еще не достаточно ясно для свободно живущих ресничных и кольчатых червей, но для других животных, размножающихся бесполым путем, в особенности путем деления, это несомненно.
Все эти животные главным образом — сидячие и притом обычно колониальные, в которых отдельная особь играет лишь подчиненную роль (полипообразные формы кишечнополостных, мшанки, асцидии), и не отличается активностью. Устраняется ли при этом вполне явление старости, сказать трудно. В некоторых случаях бесполое размножение делением может, повидимому, продолжаться безгранично. В других случаях, главным образом при почковании, наблюдается вполне определенное старение отдельных членов колонии, как это ясно видно в колониях мшанок и асцидий, где более старые особи постепенно отмирают. Имеются, однако, указания и на старческое вырождение целых-колоний гидроидов (Гартлауб; 1915).
Во многих случаях бесполого размножения, следовательно, ясно видна связь между жизнью индивидуума и размножением. Менее ясна эта связь при половом размножении. Этот способ размножения остается в качестве единственно возможного при высшем развитии индивидуальности, точно приспособленной к определенным условиям существования. Под высоко развитой индивидуальностью мы разумеем именно ту ее форму, которая характеризуется наибольшей активностью. Здесь, у высоко дифференцированных подвижных организмов, форма и величина фиксируются наиболее точно.
Продолжительность жизни особи определяется здесь в конце концов рядом биологических факторов и в особенности условиями размножения. По Вейсманну, жизнь дана каждому виду возможно более короткая, насколько это возможно при данном способе {80} размножения, т.-е. при данных условиях плодовитости и выживания молоди вплоть до достижения ею половозрелого состояния. В некоторых случаях животные размножаются однократно, и тогда они гибнут тотчас же после воспроизведения потомства.
Резюмируя сказанное, мы можем отметить, что в ряду животных осуществляется процесс развития все более совершенных и стойких форм индивидуальности. По биологическим условиям только у сидячих форм возможно существование высших индивидуальностей (колоний) с неограниченным ростом. В этом случае иногда еще можно, как и у простейших, говорить о их бессмертии. У свободно живущих форм рост ограниченный. У низших из них иногда еще наблюдается периодическое появление зон роста, приводящих к бесполому размножению. У высших форм точная определенность организации исключает возможность бесполого размножения. Животные с вполне стойкой активной индивидуальностью размножаются только отдельными половыми клетками. Все остальное тело рано или поздно гибнет. Смерть есть плата за приобретение высшей индивидуальности с длительным существованием.
Все виды размножения можно рассматривать как частичное омоложение, и таким образом путем размножения организмов сохраняется непрерывность жизни.
Мы уже познакомились с некоторыми явлениями, которые можно рассматривать как омоложение всего организма. Это наблюдается иногда при значительных повреждениях, при голоде и других неблагоприятных условиях. Такое омоложение сопровождается всегда гибелью значительного числа клеток и тканей. При частичном омоложении, ведущем к регенерации, точно так же гибнет известная часть клеток и тканей. И в том и в другом случаях гибнут наиболее дифференцированные части, между тем как наименее специализированные регенеративные клетки сохраняются для восстановления утраченных частей. Этот процесс мы отмечали как дедифференцировку. Дедифференцировка простирается и на отдельные клетки, при чем и здесь она сопровождается гибелью всех дифференцированных частей протоплазмы (например, мускульных фибрилль).
Дифференцированные части организма, это как раз — те части, которые определяют его индивидуальность. Именно они фиксируют определенные соотношения с внешней средой. В особенности это {81} ясно для нервной системы. В ее организации заложен механизм, определяющий реакцию животного на те или иные воздействия внешней среды. Нервная система не только однократно отвечает на эти воздействия, но подвергается при этом и каким-то длительным изменениям, так что определенные воздействия во многих случаях определяют не только ближайшую реакцию организма, но и позднейшие его жизненные проявления. Испытанные организмом воздействия внешней среды составляют исторический базис его реакции при дальнейших воздействиях. Это явление, называемое памятью в широком смысле этого слова, составляет основное свойство-живой материи и присуще также и дифференцированной протоплазме простейших организмов. У сложных животных оно в особенности ясно выражается в функциях нервной системы. При помощи последней устанавливаются определенные соотношения организма с внешней средой и притом соотношения во многом чисто индивидуальные.
Если омоложение связано с дедифференцировкой, т.-е. с гибелью всего дифференцированного, то оно, следовательно, связано с потерей индивидуальности, если под последней понимать всю сумму индивидуального «жизненного опыта». Такой индивидуальный жизненный опыт имеется даже и у простейших, как показывают многочисленные эксперименты. И у простейших омоложение связано с гибелью дифференцированных частей тела. Не говоря уже о глубокой реорганизации, наблюдающейся при подготовке к конъюгации или при партеногенезе, при которых гибнет значительная часть ядерного аппарата и различные органеллы, даже и при простом размножении делением гибнет часть дифференцированных частей, и обновляется ядерный аппарат. Последний в особенности является здесь регулятором всех жизненных функций.
Таким образом при всех периодических процессах омоложения, как связанных с размножением, так и не связанных с ним, все характерное для индивидуума как такового перестраивается и в значительной мере гибнет. Можно прямо сказать, что индивидуумы всегда смертны, даже тогда, когда данные организмы являются потенциально бессмертными, как, например, простейшие.
Если, как мы видели, полное омоложение организма возможно только при условии полной его реорганизации за счет дедифференцированного материала, то является вопрос — не возможно ли все же {82} хотя бы некоторое продление жизни особи при условии сохранения всего характерного для данной индивидуальности, т.-е. возможно ли освежение стареющего организма без явления дедифференцировки.
Мы уже указывали, что теоретически жизнь особи даже ограниченного роста могла бы длиться очень долго при соблюдении целого ряда условий.
Прежде всего необходима такая совершенная организация, чтобы ассимиляционная деятельность даже наиболее высоко специализированных клеток как раз восполняла полностью все потери живого вещества, зависящие как от постоянной функциональной деятельности, так и от случайных повреждений. Это условие до сих пор не достигнуто ни одним организмом, и полное восстановление всех дефектов наблюдается только при избытке ассимиляции, сопровождающем рост организма. Все же у высших организмов мы наблюдаем постепенное приближение к этому идеалу и все возрастающую стойкость отдельных клеток. Однако и в наиболее совершенных организмах изнашивание не устранено, и поэтому необходима еще точная координация запасов живого вещества в отдельных органах и тканях соответственно сумме всех требований, которые к ним предъявляются в течение жизни. Соблюдение этих условий устраняет негармоничное старение организма. Очевидно эти условия заложены в самой конституции особи, и повлиять можно лишь на трату живого вещества, которая также должна быть гармоничной. Характер ассимиляционной деятельности определяется, очевидно, организацией самих клеток, в частности их ядерного аппарата, т.-е. опять-таки наследственной конституцией особи.
Однако помимо того обмен регулируется еще циркулирующими в организме гормонами. Ограничение роста, которое мы рассматривали как основную причину возникновения смерти, есть не что иное, как именно ограничение обмена. Это ограничение достигается отчасти существованием системы тормозов, имеющих, очевидно, характер гормонов. Таким образом выясняется зависимость основных жизненных процессов — роста и обмена вообще — от существования специфических гормонов, регулирующих течение этих процессов. Секреция этих гормонов зависит от состава и строения соответствующих органов, от характера протекающего в них обмена, т.-е в конце концов опять-таки определяется тем, что составляет конституцию особи.
Гормоны выделяются различнейшими, можно сказать всеми, органами и тканями, однако в большинстве случаев они ближе не {83} изучены вследствие многих трудностей, стоящих на пути такого изучения. Только у позвоночных животных, у которых имеются специальные инкреторные органы, которые никакой иной функции кроме выделения гормонов не несут, задача становится несколько более доступной. Так как каждый такой орган характеризуется своим специфическим обменом, то является возможность тем или иным способом повлиять на его функцию. Таким образом изучение гормонов, регулирующих обмен, изучение функции органов, выделяющих эти гормоны, дает нам возможность овладеть до известной степени этой функцией, а следовательно и надежду вмешаться в деятельность механизма, регулирующего явления обмена и роста особи.
Устранением чрезмерного действия тормозов ассимиляции, быть может, удалось бы до известной степени устранить последствия несовершенного восстановления живого вещества и возникающих отсюда нарушений обмена, накопление продуктов распада и старение организма. Все это, конечно, лишь до известной степени, так как и сами органы внутренней секреции изнашиваются вследствие автономного ограничения ассимиляции, и поддержка их функции возможна, очевидно, лишь в известных пределах, определяемых опять-таки конституцией особи.
Попытки добиться омоложения организма действием гормонов начинаются с известного эксперимента Броун-Секара, произведенного им над самим собой в 1889 году. Опыт состоял в подкожном впрыскивании экстракта, полученного из семенника млекопитающих. Последовавшие затем другие эксперименты не дали тех результатов, на которые рассчитывал Броун-Секар, и вопрос на некоторое время заглох.
В новейшее время такого рода попытки были возобновлены Гармсом, который применил к старым млекопитающим метод трансплантации кусков семенников молодых животных. Этот автор получил блестящие результаты, при чем кроме восстановления угасшей половой способности наблюдалось и полное восстановление жизненных сил вообще. В меньшей мере, однако, омоложение касается центральной нервной системы и органов чувств.
К этим опытам тесно примыкают эксперименты Штейнаха, который применил также и иной метод. Штейнах вместо пересадки половых желез возбуждал их к новой усиленной деятельности путем перевязки у самцов семенного канатика. При этом наблюдаются сначала дегенерация половых клеток и разрастание {84} промежуточной ткани, а затем и регенерация половой ткани. При этом также получался значительный эффект в смысле повышения жизнедеятельности и даже заметного продления жизни. У самок омоложение достигалось пересадкой кусков яичников, взятых от молодых животных. Во всех этих случаях восстановление половой способности сопровождалось общим подъемом жизненных сил, а также и временным устранением целого ряда других старческих явлений, как, например, восстановлением волосяного покрова и др.
Достигнутое при этом омоложение представляет результат воздействия гормонов, выделяемых инкреторными частями половых желез. Весьма вероятно, что это омоложение окажется в значительной мере односторонним. Внимание исследователей остановилось прежде всего на половой железе, так как угасание ее деятельности представляет собой одно из наиболее ярких и ранних проявлений старости.
Однако обмен веществ, к изменениям которого, как мы видели, главным образом и сводится сущность старческих изменений организма, регулируется деятельностью целого ряда желез внутренней секреции. Так, например, не только половые железы усиливают обмен, но в особенности щитовидная железа выделяет гормон, весьма значительно повышающий обмен. Она же играет известную роль при синтезе высших белков. В некоторых отношениях обменом ведают и надпочечники и гипофиза. Кроме этого, эндокринные железы влияют еще на целый ряд других органов, регулируя их обмен, рост и развитие. Так, например, щитовидная железа оказывает влияние на рост костей и кожи, надпочечники — на построение нервной и мышечной ткани, на распределение пигментов и на рост волос. Гипофиза влияет на построение и рост костей и, кроме того, регулирует деятельность различных желез. Кроме того инкреторные железы регулируют формообразование животного и поддерживают уже сложившиеся формы. И в этом отношении, как и в других, обнаруживается также взаимная зависимость функций отдельных желез, которые частью возбуждают друг друга (как, например, щитовидная и половая или половая и надпочечники), частью взаимно тормозят (как зобная и половая или щитовидная и гипофиза).
Наконец, наблюдается и теснейшая взаимная связь функций этих желез с деятельностью нервной системы. Известно, например, воздействие гормонов половых желез на деятельность нервной системы и даже на психику животных, и не менее известно возбуждающее действие гормона надпочечников (адреналина) на симпатическую нервную систему. {85}
Получается очень сложная цепь зависимостей, в которой пока еще разобраться невозможно.
Мы еще не знаем, какие органы играют действительно руководящую роль в тех изменениях обмена, которые идут впереди других старческих изменений. Дальнейшие исследования желез внутренней секреции должны нам указать, нет ли более надежных путей для борьбы с преждевременной старостью, чем тот путь, на который стали Гармс и Штейнах.
Вейсманн рассматривал смерть как вторичное приобретение сложных животных, как полезное приспособление их в борьбе за существование. Многие другие авторы считают смерть неизбежной, конечной фазой жизненного цикла всех организмов, т.-е. первичным свойством живой материи. Обе стороны до известной степени правы. Простейшие действительно потенциально бессмертны, так как они путем непрерывного размножения, связанного с обновлением всей организации (иногда чередующегося и с еще более глубокой реорганизацией ядерного аппарата при партеногенезе или типичном половом процессе), избегают старости и смерти.
Организм простейших также изнашивается и, несомненно, даже быстрее, чем клетки сложного организма, но это изнашивание не успевает привести к смерти, так как особи обладают слишком кратковременным существованием. Жизнь особи прерывается иным путем, и только потому у них нет смерти.
Сложные животные лишь в редких случаях (чаще сидячие формы) обладают непрерывным ростом и подобно простейшим, достигнув предельного размера, разделяются на два новых индивидуума. В этом случае замечается подобное же обновление всей организации, как и у простейших, и вегетативное размножение возможно, быть может, неограниченно долго.
Обычно же рост является ограниченным, и в особенности точно фиксированной является форма животного, точно приспособленная к определенным биологическим условиям (особенно у свободно перепевающихся животных). Так как вегетативное размножение делением или почкованием связано хотя бы и с временным нарушением формы животного, то у высших животных оно оказывается невозможным.
Ограничение ассимиляции и прекращение клеточных делений, связанное с ограничением роста, лишает животное того средства {86} обновления организации, которое и делает простейших бессмертными. Сложные животные смертны благодаря ограничению их роста. С другой стороны, стойкость такого высшего организма несомненно возрастает. Если смерть и есть в конце концов результат изнашивания организма вследствие несовершенного его восстановления, то у нас нет никаких оснований думать, что у простейших это восстановление во время индивидуальной жизни совершеннее, чем у сложных животных.
Ограничение роста сложных животных осуществляется иногда фиксированием числа клеточных делений, а следовательно и общего числа клеток всего организма. У многих животных это число оказывается действительно строго определенным. В этом случае, однако, весь организм естественно изнашивается еда сравнительно быстро. В других случаях фиксированным оказывается число клеток лишь в некоторых наиболее дифференцированных органах, как, например, в нервной системе. У таких животных определенность формы поддерживается не столько благодаря точному фиксированию числа клеточных делений, сколько благодаря высокой регуляторной способности организма. В этом случае возможна и борьба со старостью путем постепенной замены изношенных клеток свежими. Такая смена возможна, однако, лишь в тех органах, в которых не необходима безусловная цельность всей структуры в течение всей жизни, т.-е. непрерывность определенной связи всех его клеток. Поэтому нервная система не может быть обновляема, и в связи с этим, повидимому, смерть высших животных определяется именно изнашиванием нервной системы. Увеличение общей массы последней, и, следовательно, распределение работы на большее число клеток ведет, очевидно, к увеличению продолжительности индивидуальной жизни („фактор цефализации”).
Во всяком случае продолжительность жизни определяется всей организацией животного и не менее характерна для данного вида или формы, чем любые морфологические или физиологические признаки. Полиморфизм возможен и в отношении длительности жизни. Как и вся организация, и продолжительность жизни определяется в конце концов структурой половых клеток, составом его „генов”. При этом одним из условий максимального долголетия является строгая гармоничность всей организации. Омоложение состарившихся уже частей, а также и целого организма в некоторых случаях вполне возможно. При поранениях, а иногда и под влиянием голода или иных неблагоприятных условий, наблюдается {87} дедифференцировка поврежденных частей, а иногда и всего организма. Все высоко специализированные части гибнут, если они значительно повреждены. За счет оставшихся мало дифференцированных частей образуются ткани, до известной степени эмбрионального характера, клетки которых вновь приобрели способность размножаться путем деления. Таким образом восстанавливается утраченная часть, или иногда даже весь организм, являющийся в этих случаях явно омоложенным.
Однако, как сказано, наиболее дифференцированные части гибнут, и таким образом высоко специализированные органы, как нервная система, не могут быть реорганизованы, а восстанавливаются заново за счет менее дифференцированных тканей (нервная система, например, из эпидермиса). Так как все характеризующее индивидуум как таковой определяется его дифференцированными частями, в частности нервной системой, то, очевидно, омоложение организма путем его реституции связано с потерей всего жизненного опыта и вообще всего того, что выделяет данный организм из среды ему подобных. Омоложение путем реституции связано с гибелью индивидуальности. Бесполое и половое воспроизведение как частные случаи такого омоложения, конечно, уже явно ведут к образованию совершенно новых индивидуальностей.
Таким образом существование индивидуумов всегда ограничено во времени. Если не смерть, то иные причины ведут к потере индивидуальности.
Наименее стойка индивидуальность у простейших, которые в благоприятных условиях обладают крайне кратковременным существованием, так как нарастание живого вещества уже очень быстро достигает своего предела, когда оно приводит к размножению. При неблагоприятных условиях простейшие могут избегнуть гибели путем энцистирования, которое также связано с утерей дифференцированных частей, т.-е. с утратой индивидуальности.
В колониальных формах простейших появляется впервые высшая, более стойкая, индивидуальность, при чем жизнь всей колонии определяет жизнь Отдельных особей. Естественно, что таким образом отдельные особи приобретают более длительное существование.
Еще более устойчивым является организм сложных животных благодаря совершенному разделению труда между отдельными его элементами. В этом случае и отдельные клетки, попадающие в строго определенные условия существования и обладающие соответственно {88} своей специализации не менее определенным обменом, могут в совершенстве приспособиться к выполнению своих функций в течение очень долгого времени.
Мы пришли к выводу, что у всех сложных животных имеется некоторый внутренний аппарат, устанавливающий предел роста и передаваемый по наследству так же, как и прочие признаки организации. Этот аппарат ограничивает в то же время и жизнь определенными нормами, так как ограничение роста есть ограничение ассимиляционной деятельности, ведущее к ограничению регенерационной и вообще регуляторной способности, без которых невозможно совершенное восстановление функциональных повреждений. Несовершенное восстановление мы обозначаем как изнашивание. Существенным результатом этого изнашивания являются падение обмена ниже нормы и неполное удаление продуктов распада как первые признаки старческих изменений, за которыми следуют и все другие.
При настоящем изложении мы только констатировали факт изнашивания живого вещества, которое ясно наблюдается в том случае, если его запас является строго определенным, т.-е. если рост ограничен известными пределами, и поэтому связываем явления, старости и смерти с ограничением роста.
Такое объяснение оставляет нас до некоторой степени неудовлетворенными. Является вопрос, почему же невозможно-совсем избегнуть этого изнашивания? Ведь все же способность самовосстановления есть одно из наиболее характерных свойств живого вещества. Однако в полной мере, как мы видели, это справедливо лишь по отношению к недифференцированному живому веществу, которое вне организма способно к неограниченному самовосстановлению. В различных бессмертных культурах всегда сохраняется известный запас недифференцированного живого вещества, и относительное количество его не уменьшается. В живом организме, повидимому, всегда происходит обратное. Исследования Ружички показали, что живое вещество подвергается в организме постепенному уплотнению. Протоплазма испытывает возрастные изменения, и эти изменения заставляют организм различно реагировать на одни и те же воздействия (рис. 16). Мало дифференцированные ткани несомненно испытывают возрастные изменения под тормозящим влиянием самого организма, но дифференцированные ткани и вне организма не способны к полному самовосстановлению, и мы предполагали существование внутриклеточного аппарата, {89} ограничивающего ассимиляцию и рост в виде, например, ограниченного запаса некоторых деятельных веществ (энзимов).
Ближе мы пока не можем ответить на вопрос о сущности возрастных изменений; мы вынуждены, следовательно, лишь констатировать, что ткани, обладающие ограниченным запасом живого вещества, всегда стареют. В качестве гипотезы можно, однако, высказать еще следующие соображения. Мы знаем, что функциональные воздействия ускоряют дифференцировку развивающихся тканей и органов. Мы знаем также, что всякое воздействие на живое вещество оставляет известный след, вызывая какое-то физико-химическое изменение, являющееся исторической основой его дальнейших индивидуальных реакций (память). Можно предположить, что не только во время развития организма, но и в течение дальнейшей жизни всякое воздействие на живое вещество, на клетку, вызывает ее ответную реакцию, выражающуюся в образовании некоторых специфических продуктов (обладающих, очевидно, несколько большей плотностью). Этот процесс, продолжающийся в течение всей жизни, нормально необратим, в особенности в тех органах, которые специализируются для выработки индивидуальной ответной реакции (нервная система). Происходит постепенное нарастание специфических продуктов за счет активной протоплазмы. Масса способного к самовосстановлению живого вещества неуклонно падает. Это и есть процесс старения. Очевидно, что течение этого процесса определяется собственно не временем, а количеством и интенсивностью воздействий на единицу массы активного живого вещества, вызвавших ответную реакцию, т.-е. количеством переживаний.
Действительное омоложение, конечно, возможно, как и раньше говорилось, при обращении процесса, ценою уничтожения всех индивидуализированных продуктов протоплазмы, т.-е. всего жизненного опыта особи.
Таким образом все же ограничение роста и образование стойких организмов с ярко выраженной индивидуальностью ведет с неизбежностью к ограничению жизни известными пределами.
Значительное расхождение между окончанием роста и концом жизни возможно лишь в наиболее совершенных организмах, при идеальной гармоничности строения с высокой дифференцировкой частей и весьма совершенным разделением труда, при которых сама возможность функциональных повреждений сводится к минимуму. Высшие животные идут по этому пути, и невозможно заранее указать {90} высшего предела возможных достижений там, где увеличение жизненного срока диктуется биологическими условиями. Разделение труда и специализация и есть тот путь, который ведет ко все более совершенному клеточному обмену и ко все более точной внутренной регуляции. Таким образом не должно удивлять, что как раз наиболее специализированные клетки обладают наиболее длительным существованием.
В самом деле жизнь животных всегда ограничена определенным сроком. Этот срок мог бы быть значительно увеличен, и если этого нет, то препятствием тому являются не общие свойства живого вещества, а биологические условия существования организмов, которые в конечном счете определяют как общую организацию животных, так и их конституцию, с которой, как мы видим, связана и определенность жизненного срока. Вейсманн отметил, что жизнь животным дана возможно более короткая — настолько, насколько это нужно, чтобы произвести на свет потомство, достаточное для поддержания существования вида. Очевидно только таким путем обезпечивается правильная смена поколений, являющаяся безусловно необходимой при эволюционном процессе. Только неспециализированные половые элементы, а отчасти и развивающийся организм, являются настолько пластичными, чтобы отвечать на раздражения внешней среды заметными изменениями, которые могут путем суммирования в ряду поколений привести к образованию новых форм. И только быстрая смена поколений может обеспечить быстрый темп такой эволюции. „Природа обладала всеми средствами сделать индивидуум бессмертным, но она выбрала для него смерть. Вместо постоянного молодения отдельных органов — посредством молодения их клеток — она выбрала молодение всего организма помощью одной клетки. Она отняла у нас бессмертие и взамен его дала нам любовь” (Е. Шульц).
Человек обладает относительно большей продолжительностью жизни, чем другие млекопитающие. И здесь, следовательно, высокое развитие индивидуальности сопровождается и значительной стойкостью организма. Полное омоложение всего организма, как мы видели, немыслимо, без утери индивидуальности, т.-е. того, что составляет всю ценность нашей жизни. Если старческое изнашивание организма неизбежно, то можно заботиться лишь о возможно равномерном изнашивании различных его частей, в особенности, нервной системы, соблюдением известных правил гигиены. Кроме того возможно еще некоторое воздействие на внутрисекреторный {91} регуляторный аппарат. Этот путь указывается в особенности явлениями дисгармоничного старения при нарушении нормальных функций этого аппарата. Во всяком случае и без того длительность жизни человека относительно большая и несомненно может быть еще заметно увеличена. Но у нас имеется еще и другое преимущество перед животными — результаты нашей творческой деятельности не гибнут вместе с нами, но накапливаются для блага будущих поколений. Так пусть же наш короткий жизненный путь освещается сознанием того, что человеческая жизнь много выше других жизней и только смерть обусловила возможность существования бессмертных творений его духа.
| {92} |
К стр. 3. По Каррелю (21), в кровяной сыворотке старых животных содержатся вещества, задерживающие рост культур.
К стр. 28. По Шмидту (22), у пчел процессы старческой дегенерации начинаются в тех отделах головного мозга, которые считаются ассоциационными центрами, и отсюда распространяются затем и на остальной мозг.
К стр. 43 и 46. И у американской плодовой мухи найдены формы с различной продолжительностью жизни, различающиеся также и некоторыми-морфологическими признаками, как, например, строением крыльев (Pearl, R. и др. 1923). Интересно, что при скрещивании долгоживущей расы дрозофилы к короткоживущей в первом поколении получены формы с несколько большей продолжительностью жизни, а во втором поколении наблюдалось явное расщепление на формы с различной продолжительностью жизни.
К стр. 56. По Эренбергу (23), проблемы бессмертия вовсе и не существует. Жизнь, это — сложный физико-химический процесс определенного направления, сопровождающийся нарастанием неживой субстанции („структур”) и ведущий только к смерти. Следовательно к смерти ведет сама жизнь и без смерти жизнь немыслима. Смерть не следствие жизни, а ее выражение. Вся точка зрения Эренберга весьма оригинальна и не может быть здесь изложена, однако я считаю нужным отметить, что Эренберг под смертью понимает и вообще прекращение существования индивидуума, т.-е. и конец индивидуальной жизни любого представителя простейших, связанный с процессом деления. Мне кажется, что при таком расширении привычное понятие смерти теряет свою определенность.
К стр. 59 и 61. Леви (Levi, с. 25) в очень обстоятельной сводке, посвященной явлениям роста, также видит в ограничении роста основное выражение возрастных изменений. По Леви, размножение клеток прекращается не вследствие причин, лежащих в самих клетках, а вследствие воздействия веществ, находящихся в питающих жидкостях организма (в кровяной сыворотке позвоночных) и меняющихся с возрастом. Однако здесь сейчас же является вопрос, почему же эти вещества с возрастом меняются. Кроме того, мне кажется, что как бы ни было велико значение гуморальных влияний, все же трудно обойтись совершенно без принятия и внутриклеточного механизма, особенно при образовании органов постоянного клеточного состава, когда число клеточных делений является строго ограниченным.
*) Жизнь — прекраснейшее изобретение природы, а смерть — ее искусственное средство, чтобы иметь много жизни.
Гете.
*) Гартманн отрицает возможность такового для Eudorina, так как е сома обладает гаплоидным числом хромозом.
1) Что накопление продуктов обмена в окружающей среде действительно оказывает вредное влияние на организмы, видно по наблюдениям Семпера над культурами Limnaeus, мельчавшими в небольших сосудах, по наблюдениям Вейсса над мальками рыб (21) и по экспериментам Мопа, Калкинса и в особенности Вудрёф и Дофлейна над культурами простейших.
1) Следует резко различать фактическую максимальную и среднюю продолжительность жизни животного в его естественной биологической среде, когда множество особей гибнет от врагов и других внешних факторов, и нормальную продолжительность жизни, до которой животное может дожить при благоприятных условиях существования.
1) Эксперименты с перевивкой саркомы крыс и карциномы мышей.
2) Нередко указывают различные исключения из этого правила. Многие такие случаи при проверке, однако, оказывались неверными. Глотка аннелид, которая регенерирует из энтодермы, оказывается, и эмбрионально развивается из энтодермы. Кишечник мшанок не только при регенерации, но и эмбрионально развивается из мезодермы. Даже у губок при изоляции клеток, по методу Вильсона, нет полной дедифференцировки, и дермальные и гастральные клетки сходятся между собой, образуя пласты соответственно своему происхождению (Гёксли).